

Voltage- and Calcium-Gated Membrane Currents Tune the Plateau Potential Properties of Multiple Neuron Types
- PMID: 37699717
- PMCID: PMC10634553
- DOI: 10.1523/JNEUROSCI.0789-23.2023
Voltage- and Calcium-Gated Membrane Currents Tune the Plateau Potential Properties of Multiple Neuron Types
Abstract
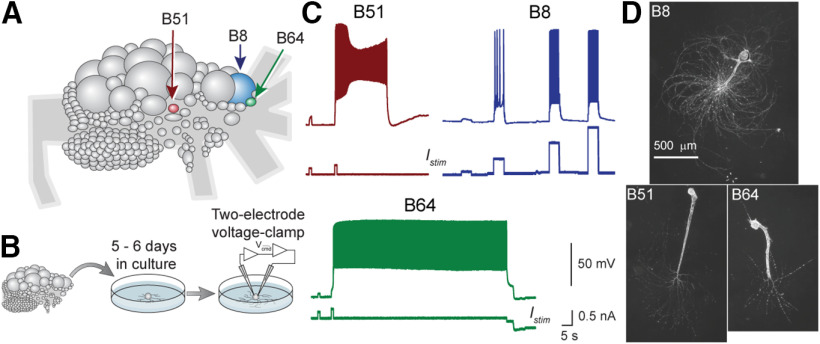
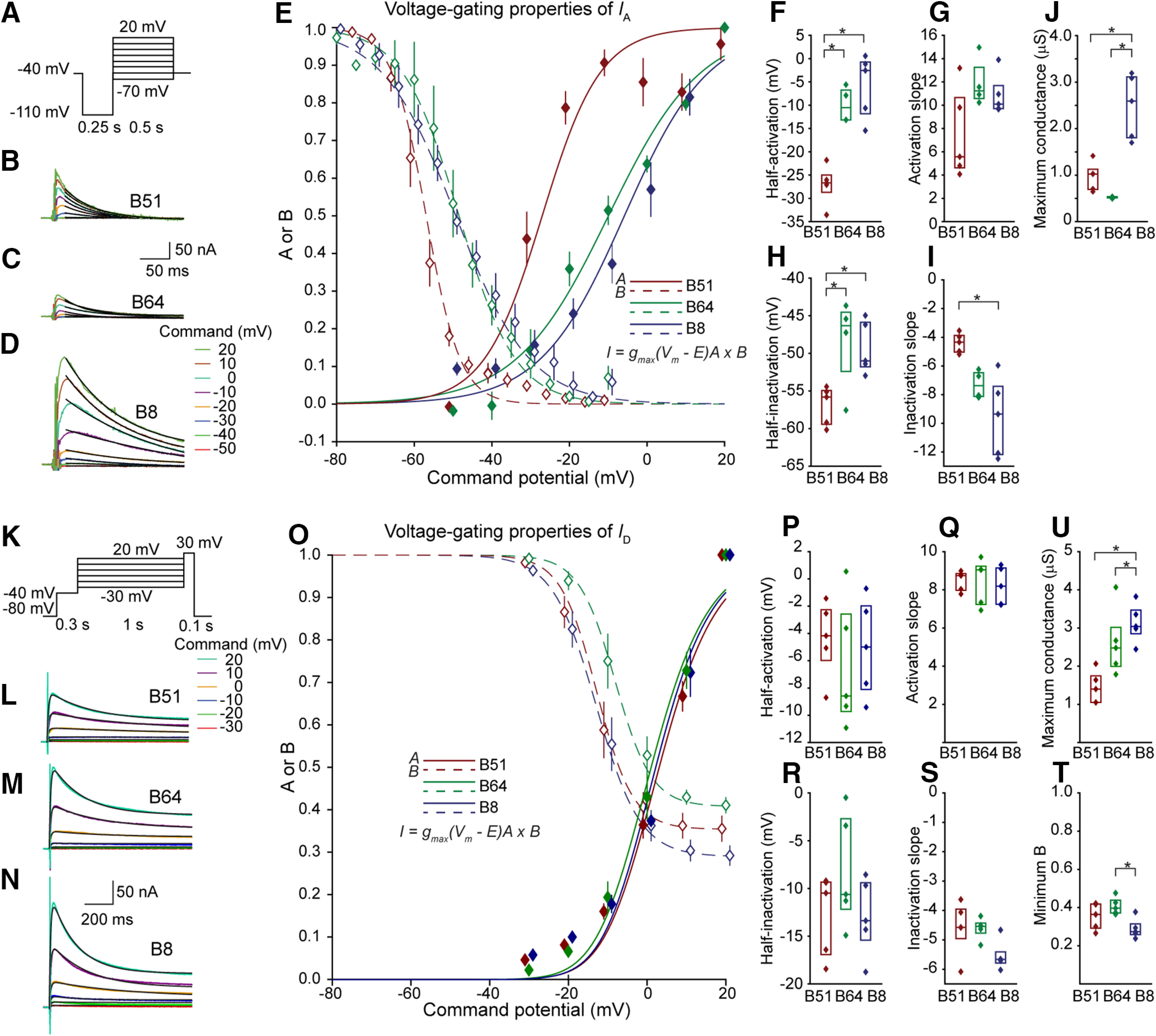
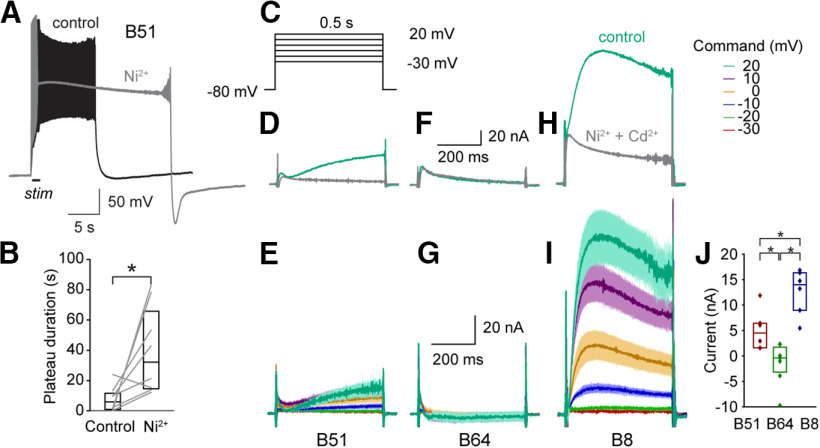
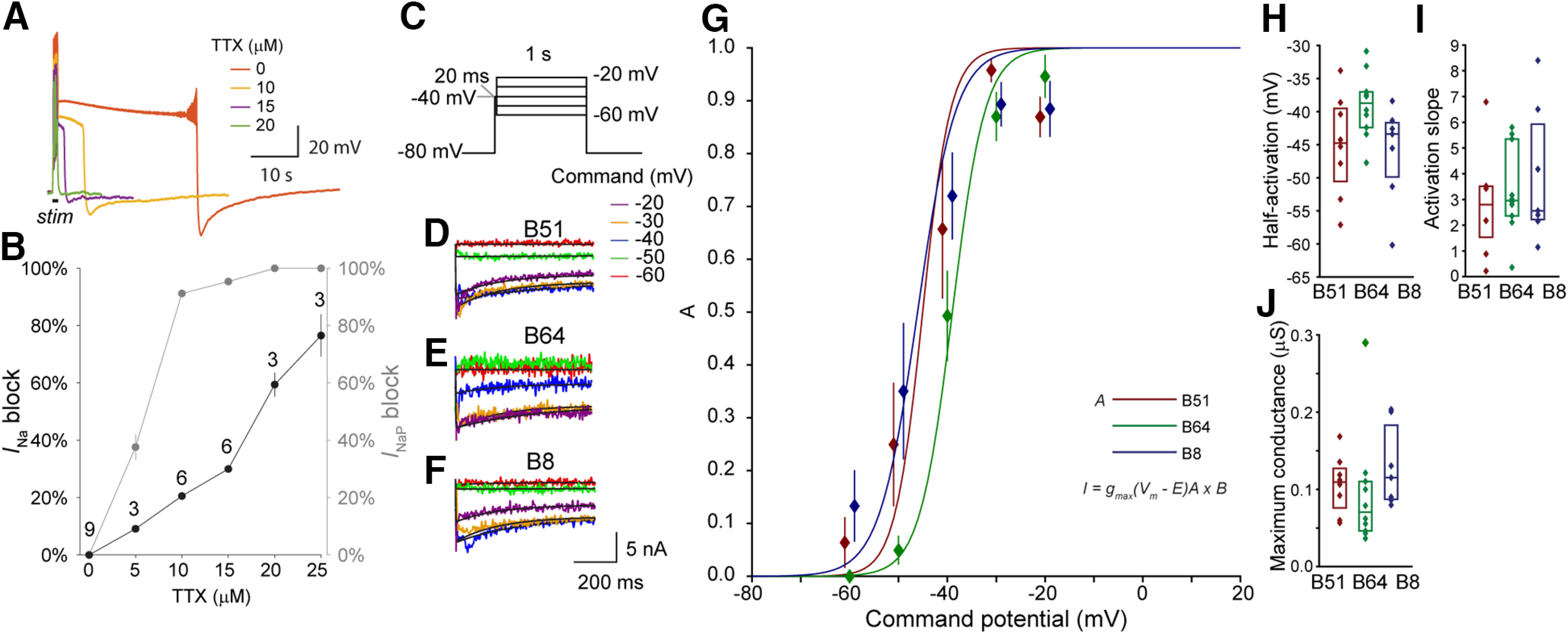
Many neurons exhibit regular firing that is limited to the duration and intensity of depolarizing stimuli. However, some neurons exhibit all-or-nothing plateau potentials that, once elicited, can lead to prolonged activity that is independent of stimulus intensity or duration. To better understand this diversity of information processing, we compared the voltage-gated and Ca2+-gated currents of three identified neurons from hermaphroditic Aplysia californica Two of these neurons, B51 and B64, generated plateau potentials and a third neuron, B8, exhibited regular firing and was incapable of generating a plateau potential. With the exception of the Ca2+-gated potassium current (I KCa), all three neuron types expressed a similar array of outward and inward currents, but with distinct voltage-dependent properties for each neuron type. Inhibiting voltage-gated Ca2+ channels with Ni+ prolonged the plateau potential, indicating I KCa is important for plateau potential termination. In contrast, inhibiting persistent Na+ (I NaP) blocked plateau potentials, empirically and in simulations. Surprisingly, the properties and level of expression of I NaP were similar in all three neurons, indicating that the presence of I NaP does not distinguish between regular-firing neurons and neurons capable of generating plateau potentials. Rather, the key distinguishing factor is the relationship between I NaP and outward currents such as the delayed outward current (I D), and I KCa We then demonstrated a technique for predicting complex physiological properties such as plateau duration, plateau amplitude, and action potential duration as a function of parameter values, by fitting a curve in parameter space and projecting the curve beyond the tested values.SIGNIFICANCE STATEMENT Plateau potentials are intrinsic properties of neurons that are important for information processing in a wide variety of nervous systems. We examined three identified neurons in Aplysia californica with different propensities to generate a plateau potential. No single conductance was found to distinguish plateau generating neurons. Instead, plateau generation depended on the ratio between persistent Na+ current (I NaP), which favored plateaus, and outward currents such as I KCa, which facilitated plateau termination. Computational models revealed a relationship between the individual currents that predicted the features of simulated plateau potentials. These results provide a more solid understanding of the conductances that mediate plateau generation.
Keywords: Aplysia; KCa; SNNAP; persistent sodium; plateau potential.
Copyright © 2023 the authors.
Figures




Similar articles
-
Divalent ion currents and the delayed potassium conductance in an Aplysia neurone.J Physiol. 1980 Jul;304:297-313. doi: 10.1113/jphysiol.1980.sp013325. J Physiol. 1980. PMID: 6255142 Free PMC article.
-
Contribution of potassium conductances to a time-dependent transition in electrical properties of a cockroach motoneuron soma.J Neurophysiol. 1999 May;81(5):2253-66. doi: 10.1152/jn.1999.81.5.2253. J Neurophysiol. 1999. PMID: 10322064
-
Intrinsic response properties of bursting neurons in the nucleus principalis trigemini of the gerbil.Neuroscience. 1998 Apr;83(3):891-904. doi: 10.1016/s0306-4522(97)00415-6. Neuroscience. 1998. PMID: 9483572
-
Differential distribution of voltage-gated channels in myelinated and unmyelinated baroreceptor afferents.Auton Neurosci. 2012 Dec 24;172(1-2):4-12. doi: 10.1016/j.autneu.2012.10.014. Epub 2012 Nov 10. Auton Neurosci. 2012. PMID: 23146622 Review.
-
The impact of single cell voltage clamp on the understanding of the cardiac ventricular action potential.Cardioscience. 1992 Sep;3(3):131-44. Cardioscience. 1992. PMID: 1384746 Review.
Cited by
-
An unusual potassium conductance protects Caenorhabditis elegans pharyngeal muscle rhythms against environmental noise.Proc Natl Acad Sci U S A. 2025 Apr 8;122(14):e2422709122. doi: 10.1073/pnas.2422709122. Epub 2025 Apr 3. Proc Natl Acad Sci U S A. 2025. PMID: 40178897 Free PMC article.
-
Multiple patterns of persistent inward currents with multiple types of repetitive firings in medullary serotonergic neurons of mice: An experimental and modeling study.PLoS Comput Biol. 2025 Apr 9;21(4):e1012918. doi: 10.1371/journal.pcbi.1012918. eCollection 2025 Apr. PLoS Comput Biol. 2025. PMID: 40203009 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous