

Metabolic heterogeneity of tissue-resident macrophages in homeostasis and during helminth infection
- PMID: 37699869
- PMCID: PMC10497597
- DOI: 10.1038/s41467-023-41353-z
Metabolic heterogeneity of tissue-resident macrophages in homeostasis and during helminth infection
Erratum in
-
Author Correction: Metabolic heterogeneity of tissue-resident macrophages in homeostasis and during helminth infection.Nat Commun. 2024 Jul 31;15(1):6452. doi: 10.1038/s41467-024-50681-7. Nat Commun. 2024. PMID: 39085251 Free PMC article. No abstract available.
Abstract
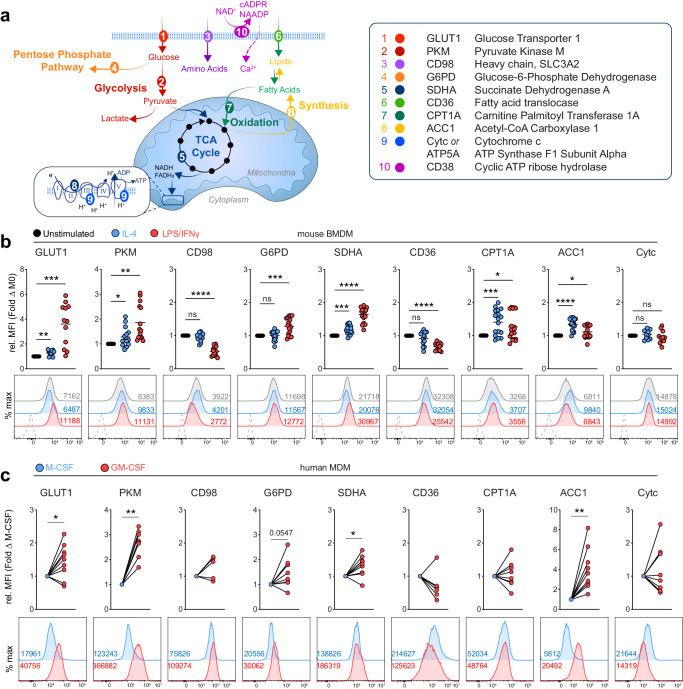
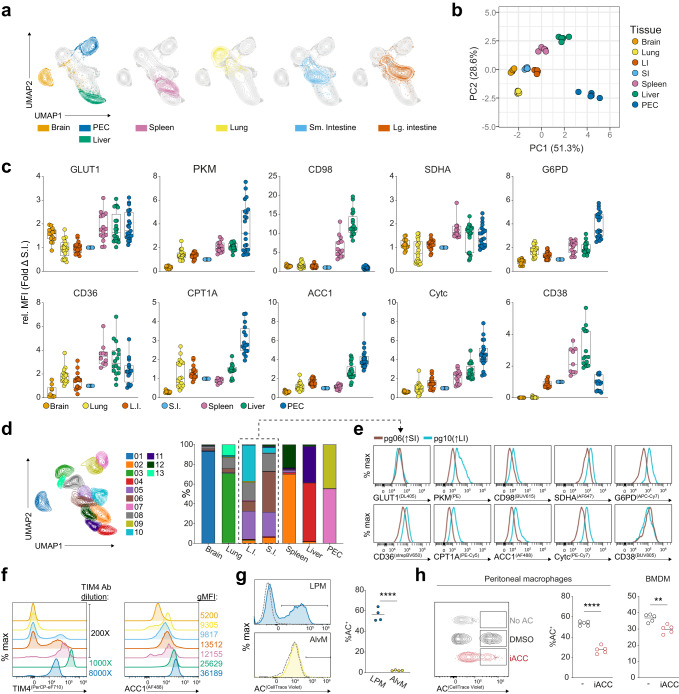
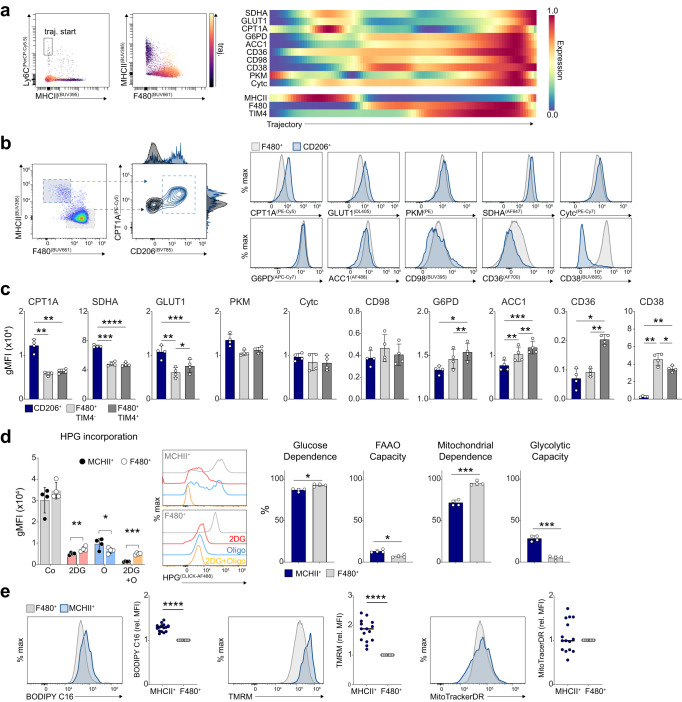
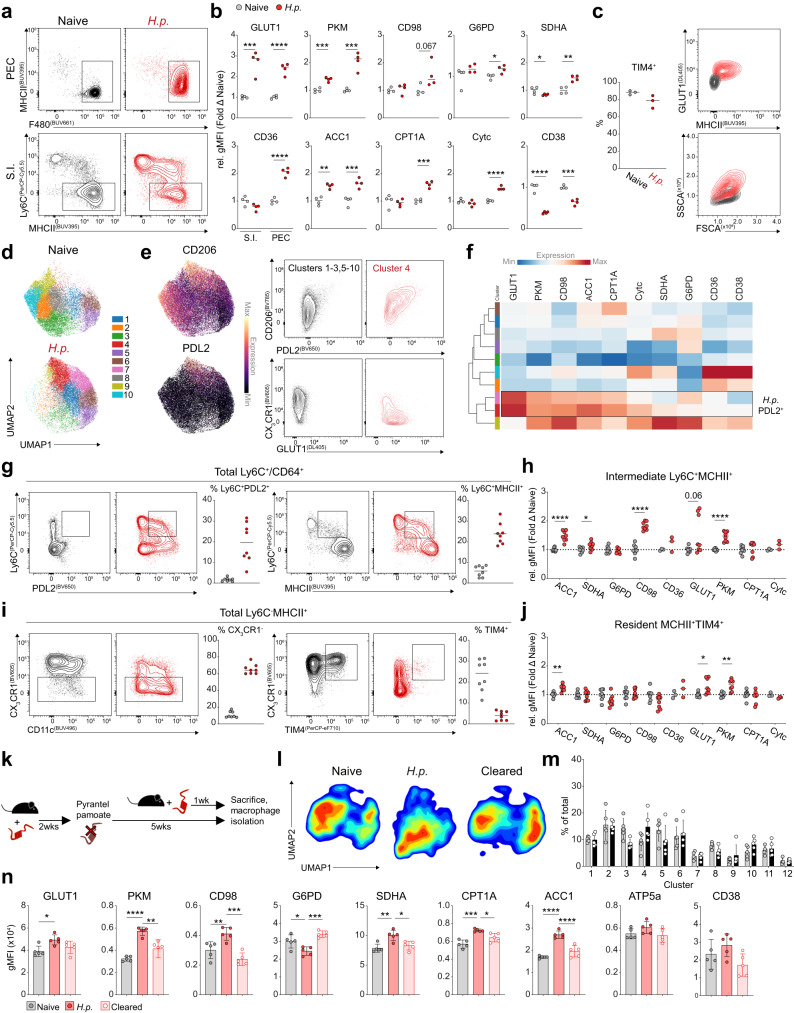
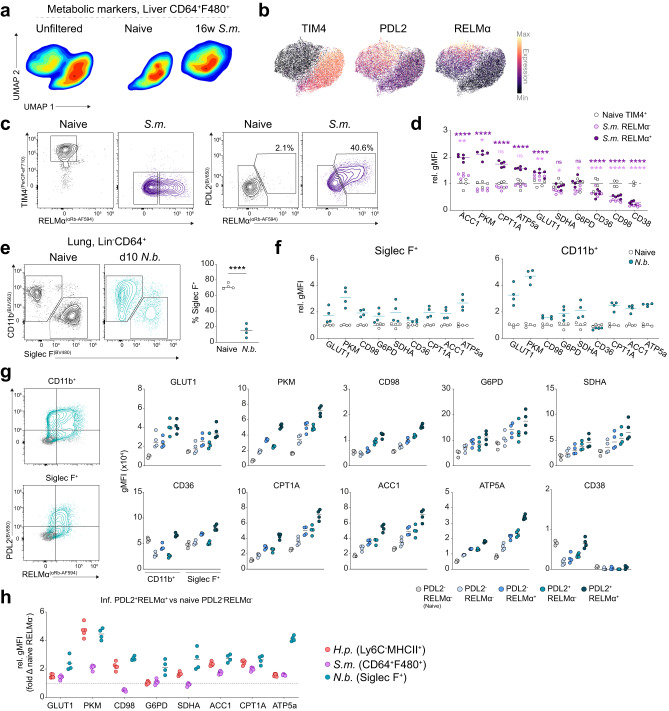
Tissue-resident macrophage populations constitute a mosaic of phenotypes, yet how their metabolic states link to the range of phenotypes and functions in vivo is still poorly defined. Here, using high-dimensional spectral flow cytometry, we observe distinct metabolic profiles between different organs and functionally link acetyl CoA carboxylase activity to efferocytotic capacity. Additionally, differences in metabolism are evident within populations from a specific site, corresponding to relative stages of macrophage maturity. Immune perturbation with intestinal helminth infection increases alternative activation and metabolic rewiring of monocyte-derived macrophage populations, while resident TIM4+ intestinal macrophages remain immunologically and metabolically hyporesponsive. Similar metabolic signatures in alternatively-activated macrophages are seen from different tissues using additional helminth models, but to different magnitudes, indicating further tissue-specific contributions to metabolic states. Thus, our high-dimensional, flow-based metabolic analyses indicates complex metabolic heterogeneity and dynamics of tissue-resident macrophage populations at homeostasis and during helminth infection.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
