

Inhibition of pro-inflammatory signaling in human primary macrophages by enhancing arginase-2 via target site blockers
- PMID: 37701067
- PMCID: PMC10494319
- DOI: 10.1016/j.omtn.2023.08.023
Inhibition of pro-inflammatory signaling in human primary macrophages by enhancing arginase-2 via target site blockers
Abstract
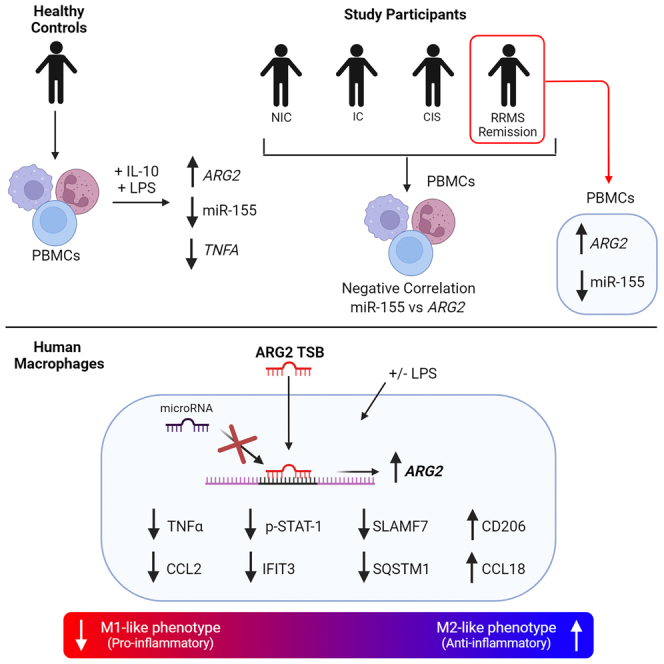
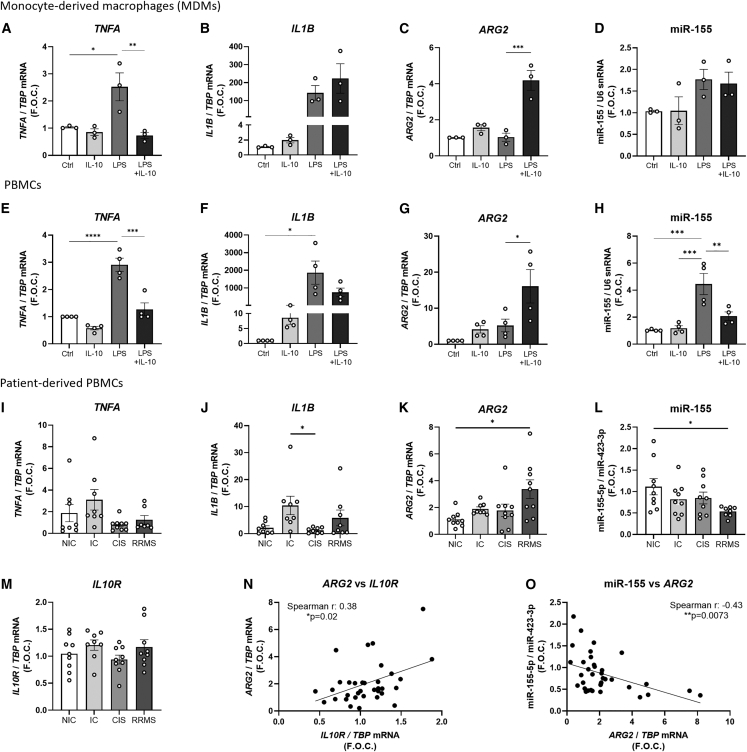
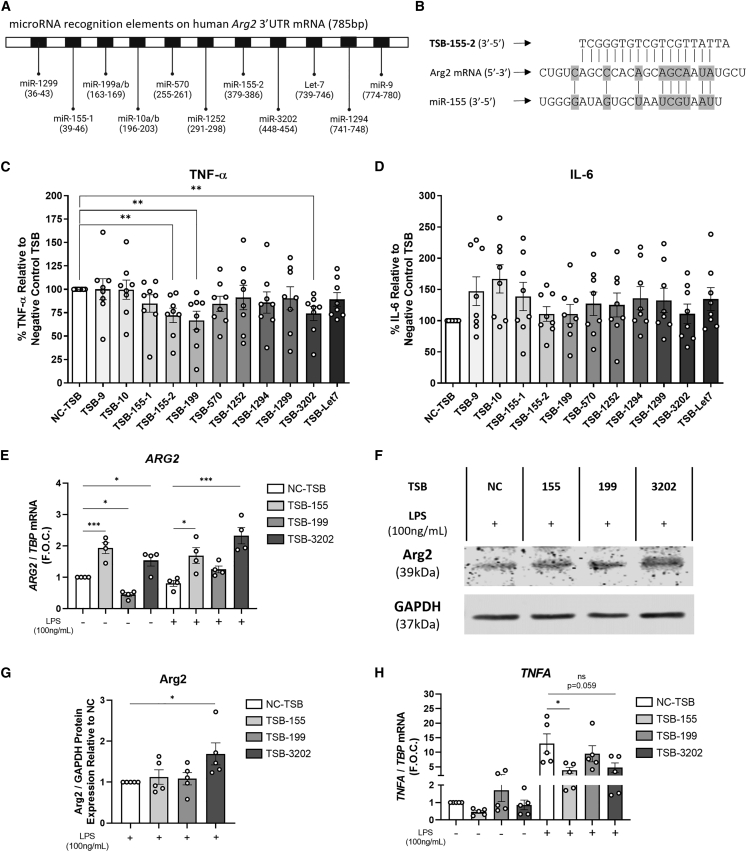
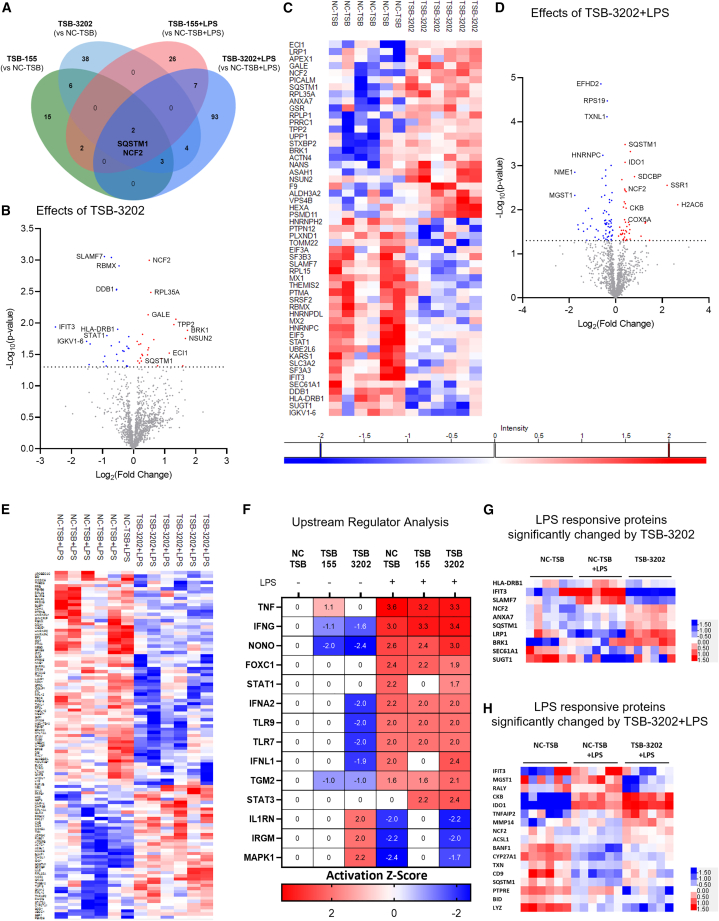
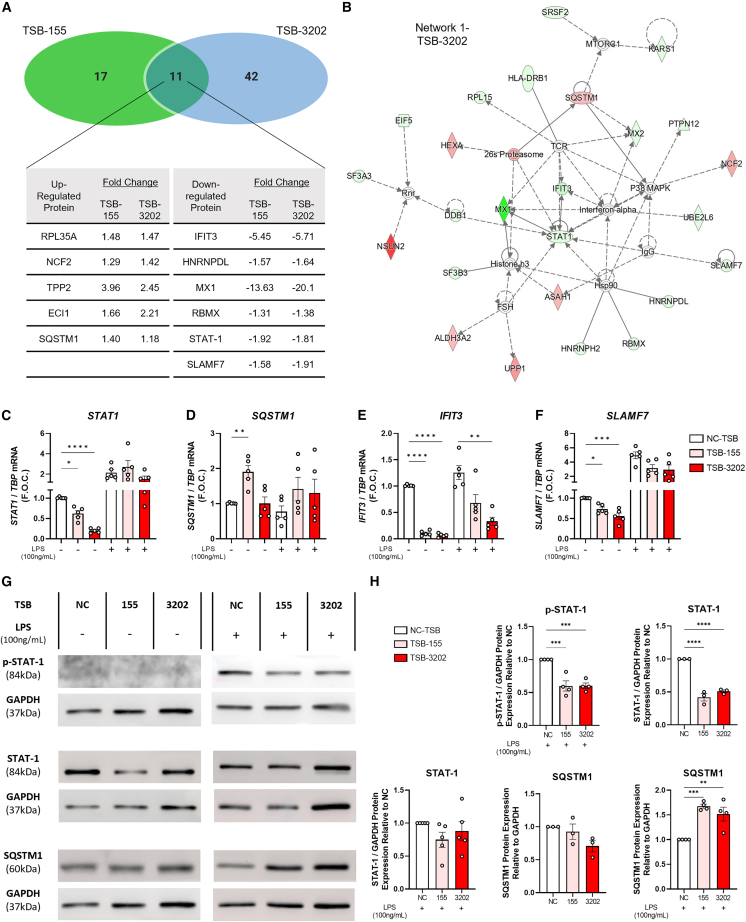
The modulation of macrophage phenotype from a pro-inflammatory to an anti-inflammatory state holds therapeutic potential in the treatment of inflammatory disease. We have previously shown that arginase-2 (Arg2), a mitochondrial enzyme, is a key regulator of the macrophage anti-inflammatory response. Here, we investigate the therapeutic potential of Arg2 enhancement via target site blockers (TSBs) in human macrophages. TSBs are locked nucleic acid antisense oligonucleotides that were specifically designed to protect specific microRNA recognition elements (MREs) in human ARG2 3' UTR mRNA. TSBs targeting miR-155 (TSB-155) and miR-3202 (TSB-3202) MREs increased ARG2 expression in human monocyte-derived macrophages. This resulted in decreased gene expression and cytokine production of TNF-α and CCL2 and, for TSB-3202, in an increase in the anti-inflammatory macrophage marker, CD206. Proteomic analysis demonstrated that a network of pro-inflammatory responsive proteins was modulated by TSBs. In silico bioinformatic analysis predicted that TSB-3202 suppressed upstream pro-inflammatory regulators including STAT-1 while enhancing anti-inflammatory associated proteins. Proteomic data were validated by confirming increased levels of sequestosome-1 and decreased levels of phosphorylated STAT-1 and STAT-1 upon TSB treatment. In conclusion, upregulation of Arg2 by TSBs inhibits pro-inflammatory signaling and is a promising novel therapeutic strategy to modulate inflammatory signaling in human macrophages.
Keywords: MT: Oligonucleotides: Therapies and Applications; arginase-2; inflammation; macrophages; miR-155; microRNA; multiple sclerosis; target site blockers.
© 2023 The Authors.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Enhancing arginase 2 expression using target site blockers as a strategy to modulate macrophage phenotype.Mol Ther Nucleic Acids. 2022 Aug 4;29:643-655. doi: 10.1016/j.omtn.2022.08.004. eCollection 2022 Sep 13. Mol Ther Nucleic Acids. 2022. PMID: 36090747 Free PMC article.
-
Precise Targeting of miRNA Sites Restores CFTR Activity in CF Bronchial Epithelial Cells.Mol Ther. 2020 Apr 8;28(4):1190-1199. doi: 10.1016/j.ymthe.2020.02.001. Epub 2020 Feb 6. Mol Ther. 2020. PMID: 32059764 Free PMC article.
-
Mitochondrial arginase-2 is essential for IL-10 metabolic reprogramming of inflammatory macrophages.Nat Commun. 2021 Mar 5;12(1):1460. doi: 10.1038/s41467-021-21617-2. Nat Commun. 2021. PMID: 33674584 Free PMC article.
-
Arginase 2 Deficiency Promotes Neuroinflammation and Pain Behaviors Following Nerve Injury in Mice.J Clin Med. 2020 Jan 22;9(2):305. doi: 10.3390/jcm9020305. J Clin Med. 2020. PMID: 31979015 Free PMC article.
-
Macrophages: The Good, the Bad, and the Gluttony.Front Immunol. 2021 Aug 12;12:708186. doi: 10.3389/fimmu.2021.708186. eCollection 2021. Front Immunol. 2021. PMID: 34456917 Free PMC article. Review.
Cited by
-
Arginine metabolism in myeloid cells in health and disease.Semin Immunopathol. 2025 Jan 25;47(1):11. doi: 10.1007/s00281-025-01038-9. Semin Immunopathol. 2025. PMID: 39863828 Free PMC article. Review.
References
-
- Schultze J.L., Schmieder A., Goerdt S. Macrophage activation in human diseases. Semin. Immunol. 2015;27:249–256. - PubMed
-
- Chu F., Shi M., Zheng C., Shen D., Zhu J., Zheng X., Cui L. The roles of macrophages and microglia in multiple sclerosis and experimental autoimmune encephalomyelitis. J. Neuroimmunol. 2018;318:1–7. - PubMed
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
