

Block-pulse integrodifference equations
- PMID: 37702828
- PMCID: PMC10500018
- DOI: 10.1007/s00285-023-01986-6
Block-pulse integrodifference equations
Abstract
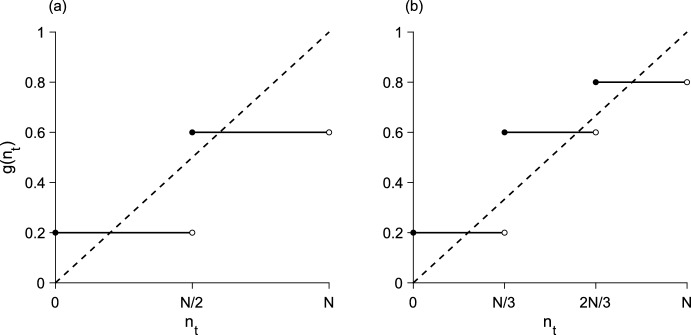
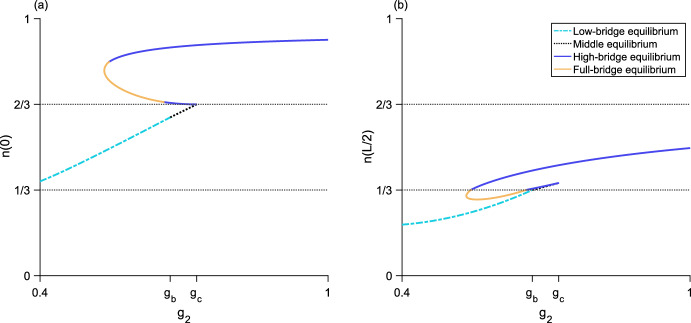
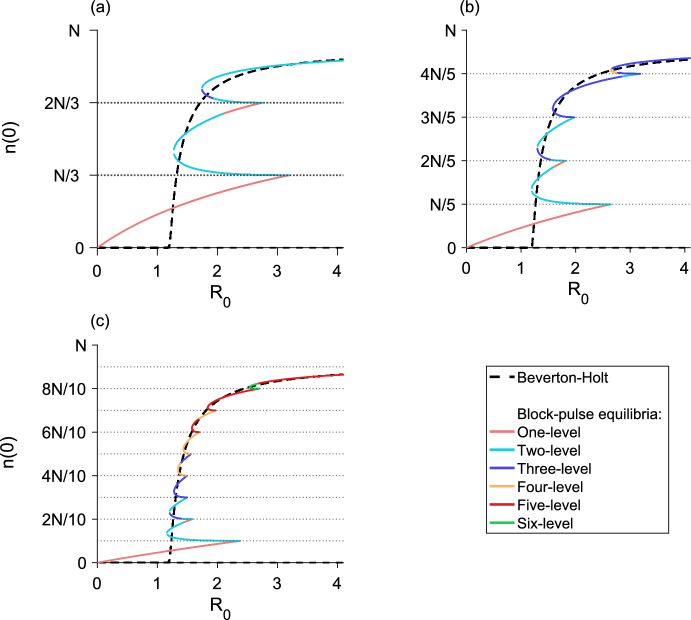
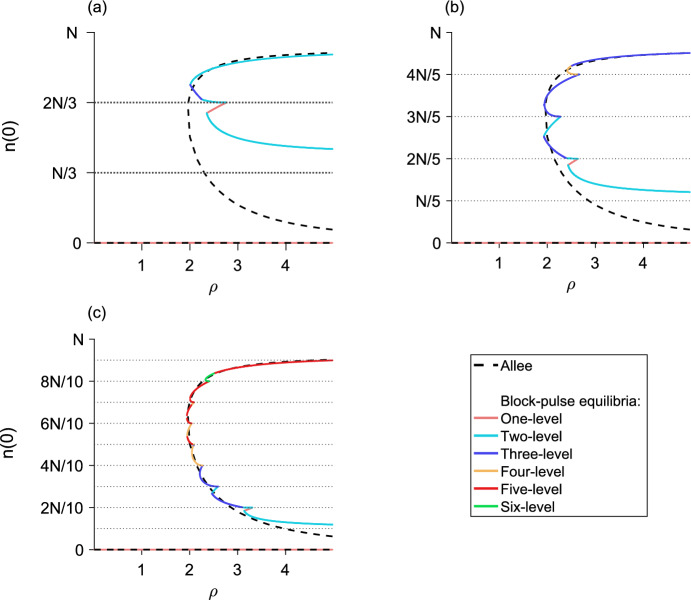
We present a hybrid method for calculating the equilibrium population-distributions of integrodifference equations (IDEs) with strictly increasing growth, for populations that are confined to a finite habitat-patch. This method is based on approximating the growth function of the IDE with a piecewise-constant function, and we call the resulting model a block-pulse IDE. We explicitly write out analytic expressions for the iterates and equilibria of the block-pulse IDEs as sums of cumulative distribution functions. We characterize the dynamics of one-, two-, and three-step block-pulse IDEs, including formal stability analyses, and we explore the bifurcation structure of these models. These simple models display rich dynamics, with numerous fold bifurcations. We then use three-, five-, and ten-step block-pulse IDEs, with a numerical root finder, to approximate models with compensatory Beverton-Holt growth and depensatory, or Allee-effect, growth. Our method provides a good approximation for the equilibrium distributions for compensatory and depensatory growth and offers numerical and analytical advantages over the original growth models.
Keywords: Allee effects; Block-pulse series; Integrodifference equations; Population dynamics; Spatial ecology.
© 2023. The Author(s).
Conflict of interest statement
The authors have no competing interests to declare that are relevant to the content of this article.
Figures













References
-
- Allee WC, Park O, Emerson AE, Park T, Schmidt KP. Principles of animal ecology. Philadelphia: Saunders Company; 1949.
-
- Andersen M. Properties of some density-dependent integrodifference equation population models. Math Biosci. 1991;104:135–157. - PubMed
-
- Apostol TM. Mathematical analysis. 2. Reading: Addison-Wesley Publishing Company; 1974.
-
- Babolian E, Masouri Z, Hatamzadeh-Varmazyar S. New direct method to solve nonlinear Volterra–Fredholm integral and integro-differential equations using operational matrix with block-pulse functions. Prog Electromagn Res B. 2008;8:59–76.
-
- Balcı MA, Sezer M. Hybrid Euler–Taylor matrix method for solving of generalized linear Fredholm integro-differential difference equations. Appl Math Comput. 2016;273:33–41.
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
