

doi: 10.1126/sciadv.adj8277.
Epub 2023 Sep 13.
CRISPR-based engineering of RNA viruses
Affiliations
- PMID: 37703376
- PMCID: PMC10499312
- DOI: 10.1126/sciadv.adj8277
Item in Clipboard
CRISPR-based engineering of RNA viruses
Sci Adv.
.
Abstract
CRISPR RNA-guided endonucleases have enabled precise editing of DNA. However, options for editing RNA remain limited. Here, we combine sequence-specific RNA cleavage by CRISPR ribonucleases with programmable RNA repair to make precise deletions and insertions in RNA. This work establishes a recombinant RNA technology with immediate applications for the facile engineering of RNA viruses.
Figures
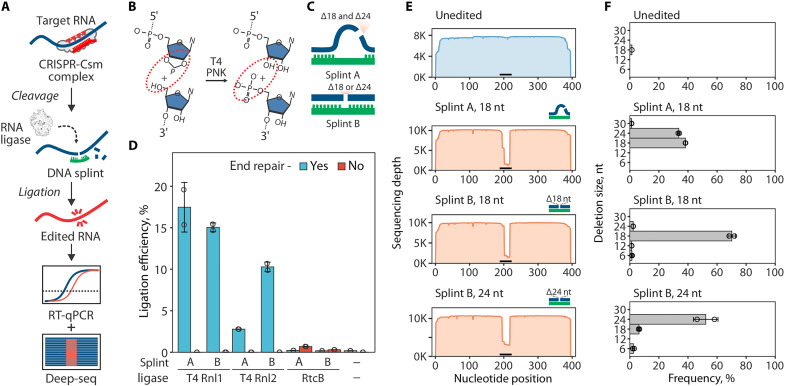
(A) Diagram of the RNA editing with type III CRISPR nucleases and splinted ligation. Type III CRISPR complexes cleave RNA in 6-nt intervals (red triangles), cutting out a portion of the target sequence. The resulting fragments are splint-ligated to introduce edits. (B) SthCsm-mediated cleavage generates 2′3′-cyclic phosphate and 5′-hydroxyl ends (left, substrate for RtcB ligase) that can be converted to 3′-hydroxyl and 5′-phosphate (right, substrate for T4 RNA ligases) using T4 PNK. (C) Two splint designs. Hybridization of the DNA splint A to complementary RNA leaves single-stranded RNA flaps (curved blue lines) that imitate a break in the tRNA anticodon loop, while splint B mimics nick in double-stranded RNA. (D) Comparison of RNA ligation efficiency with T4 RNA ligases (T4 Rnl) and RtcB ligase. Ligation efficiency was measured by performing RT-qPCR across the cut site and quantifying the signal relative to an uncut control (100%). (E) Deep sequencing of RNA ligated with splints depicted in (C). Horizontal black bar shows the target site of the SthCsm complex. (F) Quantification of editing outcomes in (E).
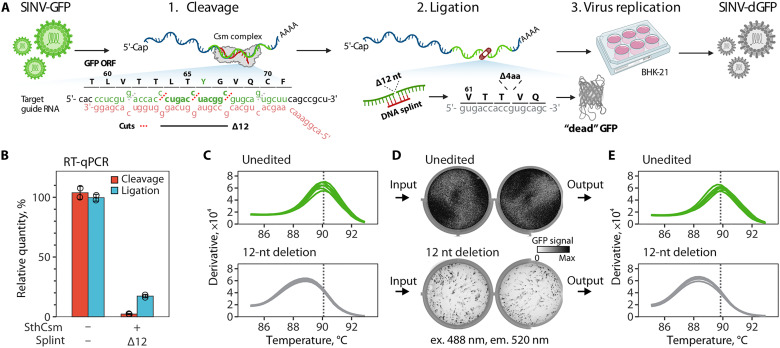
(A) Diagram showing a pipeline for deleting 12 nt from the GFP open reading frame (ORF) of the Sindbis-GFP (SINV-GFP) virus. See fig. S2A for annotated genome map and fig. S4A for additional experimental details. The deletion eliminates four codons encoding amino acids (L64, T65, Y66, and G67) forming the chromophore of the fluorescent protein, which is expected to ablate fluorescence [“dead” GFP (dGFP)]. Icons for viruses, nucleic acids, 6-well plate, and dead GFP were created with BioRender (www.biorender.com ). (B) RNA aliquots collected after the cleavage with the Csm complex (red) and ligation (blue) with T4 RNA ligase were reverse-transcribed and quantified with qPCR. Primers were designed to amplify cDNA across the target site. Relative quantities were calculated by normalizing to the uncut RNA control. (C) Melt curve analysis of the qPCR products generated with RNA after ligation in (B). Peaks indicate the melting temperature of the qPCR products. (D) BHK-21 cells were transfected with unedited or edited RNA of the SINV-GFP (two replicates each), seeded in six-well plates, and imaged 24 hours later to capture the GFP signal (excitation, 488 nm; emission, 520 nm). (E) Melt curve analysis of the RT-qPCR products that were generated with RNA extracted from supernatants of BHK-21 cells 24 hours after transfection. Primers were the same as in (B) and (C).
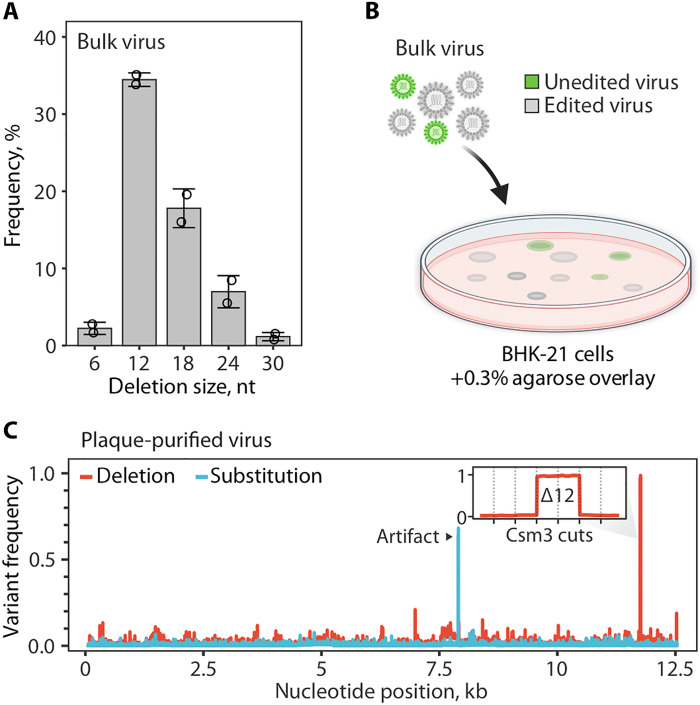
(A) Distribution of deletion sizes in sequencing reads that span the SthCsm target site in bulk edited virus. Data are shown as mean (n = 2) ± SD. Dots show individual replicates. (B) Diagram of plaque purification approach used for isolating edited virus clones. Icons for viruses and the Petri dish were created with BioRender (www.biorender.com ). (C) Frequency of nucleotide variants identified with amplicon-seq of plaque-purified virus. See fig. S7 for details on sequencing strategy and analysis of other plaque-purified viruses. Black arrow indicates a sequencing artifact associated with amplicon-seq (see fig. S7). The inset is a close-up view of the targeted genomic region. Vertical dotted lines mark predicted cut sites of the SthCsm backbone subunit (Csm3) positioned in 6-nt intervals. The 12-nt deletion was intentionally selected using a DNA splint.
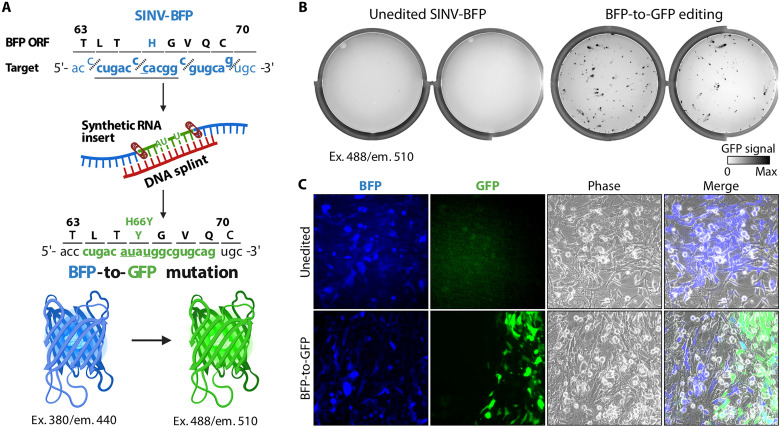
(A) Diagram showing editing strategy for substituting 12 nt (underlined) in the BFP ORF of the recombinant Sindbis-BFP (SINV-BFP) virus. The C to U substitution at 11,762 recodes histidine residue for tyrosine (H66Y), which changes the fluorescence from blue to green (BFP to GFP). Additional silent substitutions (underlined, 11,761C>A and 11,764 C>U) were included to distinguish edited RNA from the SINV-GFP used for experiments shown in Fig. 2. Icons for nucleic acids, band-aid, BFP, and GFP were created with BioRender (www.biorender.com ). (B) BHK-21 cells were transfected with the unedited or edited viral RNA (two replicates each), seeded on six-well plates, and imaged after 24-hour incubation using 488-nm excitation laser and 520-nm emission filter. This laser and filter setting does not detect BFP (excitation, 380 nm; emission, 440 nm) in the unedited control, while replication of the edited BFP-to-GFP viruses is visible. (C) Transfected BHK-21 cells shown in (B) were imaged with an inverted fluorescent microscope at ×20 magnification.
Update of
-
CRISPR-based engineering of RNA viruses.bioRxiv [Preprint]. 2023 May 20:2023.05.19.541219. doi: 10.1101/2023.05.19.541219. bioRxiv. 2023. Update in: Sci Adv. 2023 Sep 15;9(37):eadj8277. doi: 10.1126/sciadv.adj8277. PMID: 37292641 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
