

Hemin and iron increase synthesis and trigger export of xanthine oxidoreductase from hepatocytes to the circulation
- PMID: 37703667
- PMCID: PMC10506059
- DOI: 10.1016/j.redox.2023.102866
Hemin and iron increase synthesis and trigger export of xanthine oxidoreductase from hepatocytes to the circulation
Abstract
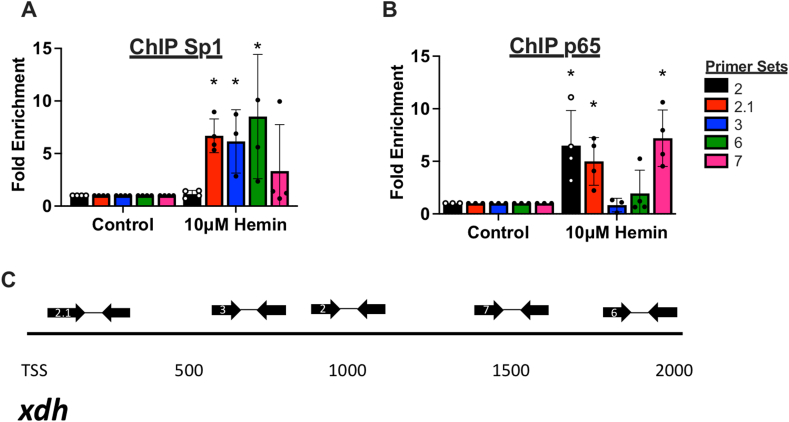
We recently reported a previously unknown salutary role for xanthine oxidoreductase (XOR) in intravascular heme overload whereby hepatocellular export of XOR to the circulation was identified as a seminal step in affording protection. However, the cellular signaling and export mechanisms underpinning this process were not identified. Here, we present novel data showing hepatocytes upregulate XOR expression/protein abundance and actively release it to the extracellular compartment following exposure to hemopexin-bound hemin, hemin or free iron. For example, murine (AML-12 cells) hepatocytes treated with hemin (10 μM) exported XOR to the medium in the absence of cell death or loss of membrane integrity (2.0 ± 1.0 vs 16 ± 9 μU/mL p < 0.0001). The path of exocytosis was found to be noncanonical as pretreatment of the hepatocytes with Vaculin-1, a lysosomal trafficking inhibitor, and not Brefeldin A inhibited XOR release and promoted intracellular XOR accumulation (84 ± 17 vs 24 ± 8 hemin vs 5 ± 3 control μU/mg). Interestingly, free iron (Fe2+ and Fe3+) induced similar upregulation and release of XOR compared to hemin. Conversely, concomitant treatment with hemin and the classic transition metal chelator DTPA (20 μM) or uric acid completely blocked XOR release (p < 0.01). Our previously published time course showed XOR release from hepatocytes likely required transcriptional upregulation. As such, we determined that both Sp1 and NF-kB were acutely activated by hemin treatment (∼2-fold > controls for both, p < 0.05) and that silencing either or TLR4 with siRNA prevented hemin-induced XOR upregulation (p < 0.01). Finally, to confirm direct action of these transcription factors on the Xdh gene, chromatin immunoprecipitation was performed indicating that hemin significantly enriched (∼5-fold) both Sp1 and NF-kB near the transcription start site. In summary, our study identified a previously unknown pathway by which XOR is upregulated via SP1/NF-kB and subsequently exported to the extracellular environment. This is, to our knowledge, the very first study to demonstrate mechanistically that XOR can be specifically targeted for export as the seminal step in a compensatory response to heme/Fe overload.
Keywords: Heme; Hemin; Hepatocytes; Iron; Oxidants; Xanthine oxidoreductase.
Copyright © 2023 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures










References
-
- Battelli M.G., Bolognesi A., Polito L. Pathophysiology of circulating xanthine oxidoreductase: new emerging roles for a multi-tasking enzyme. Biochim. Biophys. Acta. 2014;1842(9):1502–1517. - PubMed
-
- Kelley E.E. Dispelling dogma and misconceptions regarding the most pharmacologically targetable source of reactive species in inflammatory disease, xanthine oxidoreductase. Arch. Toxicol. 2015;89(8):1193–1207. - PubMed
-
- B R. Ueber dis oxidative und dis vermeindlich synthetische Bildung von Haurnsaure in Rinderleberauszug. Zeitsohr f Physiol Chem. 1905;43:497.
-
- S W. Die Ueberführung von Nucleïnbasen in Harnsäure durch die sauerstoffübertragende Wirkung von Gewebsauszügen. Arch Ges Physiol. 1899;(76):192–203.
-
- Fz S. Uber das verhalten der kulmilch gegen methylenblau und seine verwendung zur unterscheidung von ungekochter und gekochter milch. Utersuch Nahrungs- u Genussmittel. 1902;(5):1113–1121.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
