

Uncovering a mammalian neural-specific poly(A) binding protein with unique properties
- PMID: 37704377
- PMCID: PMC10546976
- DOI: 10.1101/gad.350597.123
Uncovering a mammalian neural-specific poly(A) binding protein with unique properties
Abstract
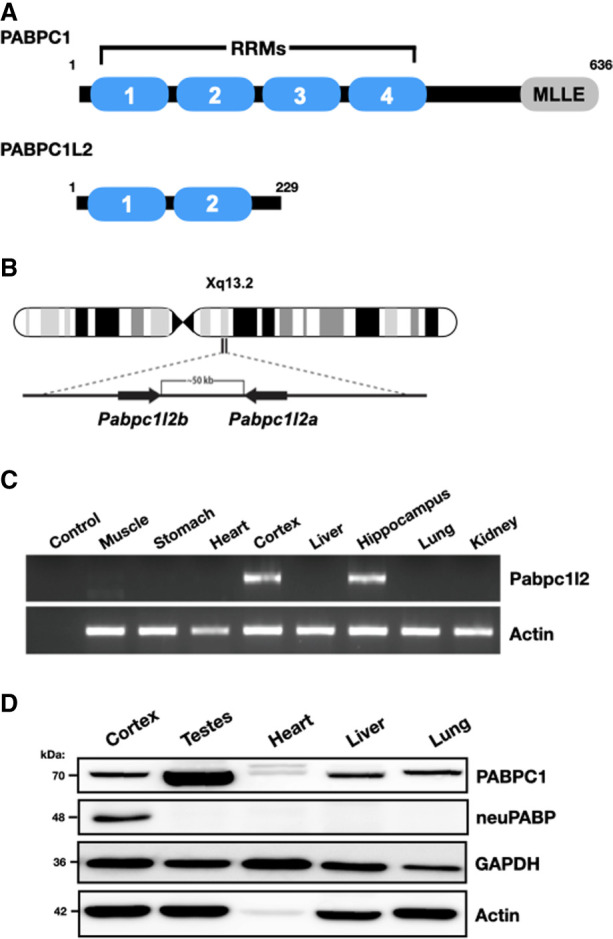
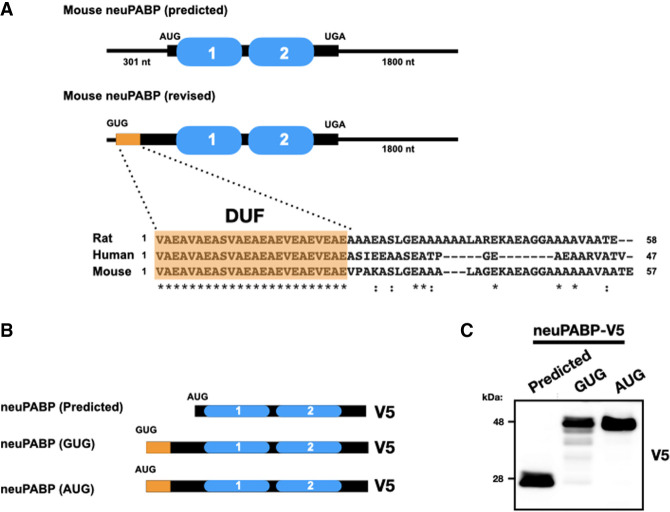
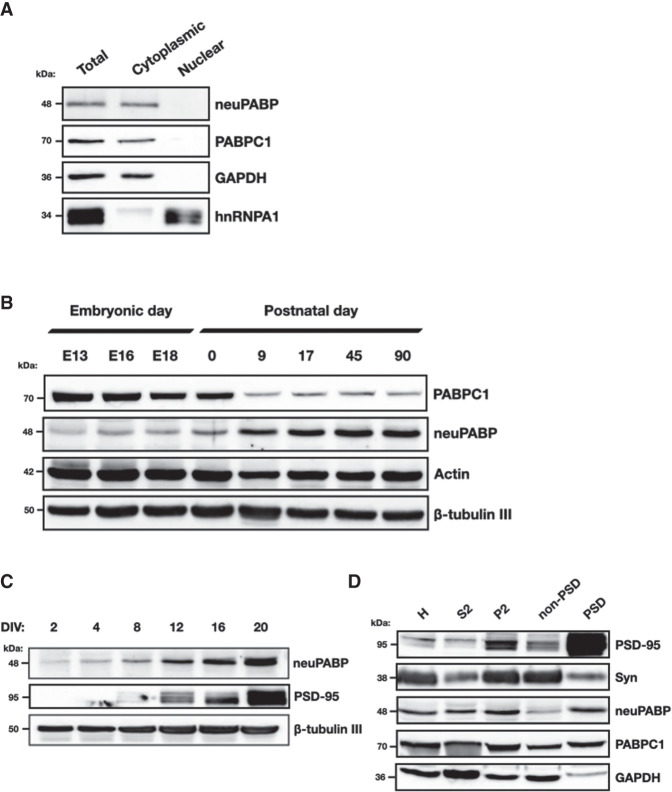
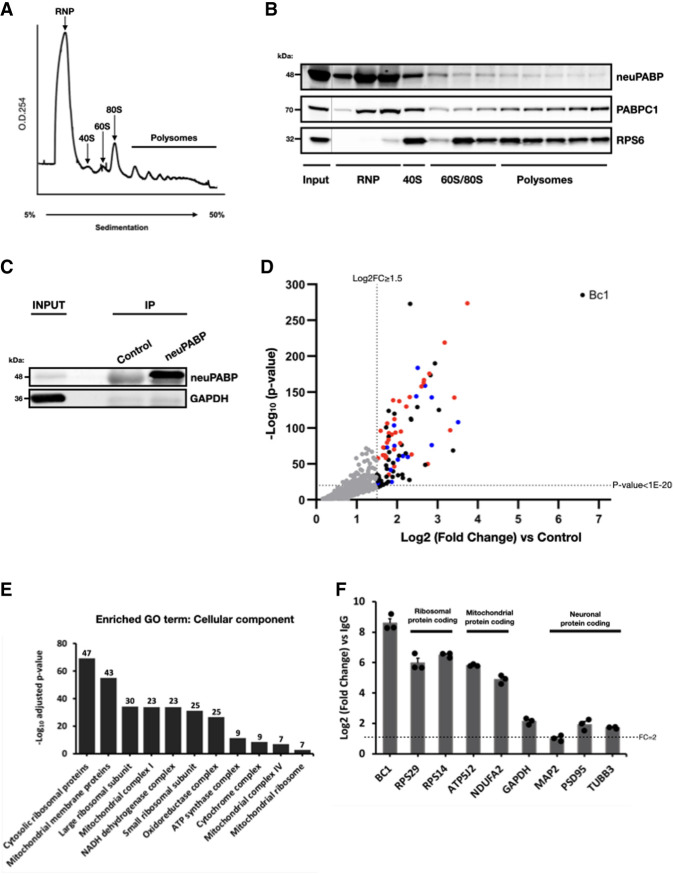
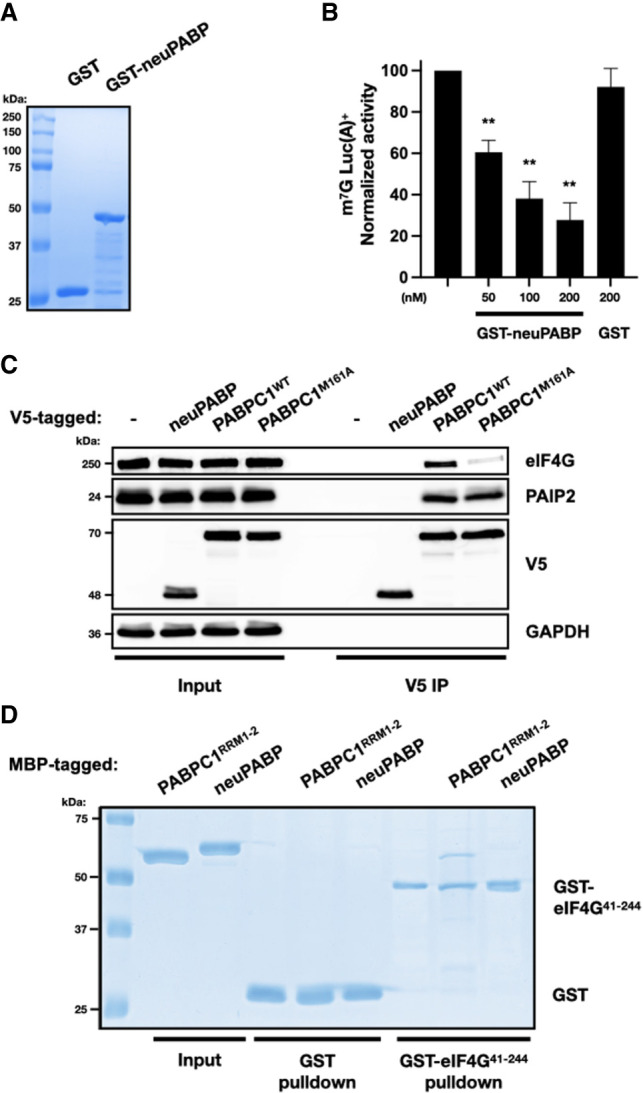
The mRNA 3' poly(A) tail plays a critical role in regulating both mRNA translation and turnover. It is bound by the cytoplasmic poly(A) binding protein (PABPC), an evolutionarily conserved protein that can interact with translation factors and mRNA decay machineries to regulate gene expression. Mammalian PABPC1, the prototypical PABPC, is expressed in most tissues and interacts with eukaryotic translation initiation factor 4G (eIF4G) to stimulate translation in specific contexts. In this study, we uncovered a new mammalian PABPC, which we named neural PABP (neuPABP), as it is predominantly expressed in the brain. neuPABP maintains a unique architecture as compared with other PABPCs, containing only two RNA recognition motifs (RRMs) and maintaining a unique N-terminal domain of unknown function. neuPABP expression is activated in neurons as they mature during synaptogenesis, where neuPABP localizes to the soma and postsynaptic densities. neuPABP interacts with the noncoding RNA BC1, as well as mRNAs coding for ribosomal and mitochondrial proteins. However, in contrast to PABPC1, neuPABP does not associate with actively translating mRNAs in the brain. In keeping with this, we show that neuPABP has evolved such that it does not bind eIF4G and as a result fails to support protein synthesis in vitro. Taken together, these results indicate that mammals have expanded their PABPC repertoire in the brain and propose that neuPABP may support the translational repression of select mRNAs.
Keywords: RNA binding protein; mRNA translation; poly(A) binding protein.
© 2023 Sharma et al.; Published by Cold Spring Harbor Laboratory Press.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases