Neuronal K+-Cl- cotransporter KCC2 as a promising drug target for epilepsy treatment
- PMID: 37704745
- PMCID: PMC10770335
- DOI: 10.1038/s41401-023-01149-9
Neuronal K+-Cl- cotransporter KCC2 as a promising drug target for epilepsy treatment
Abstract
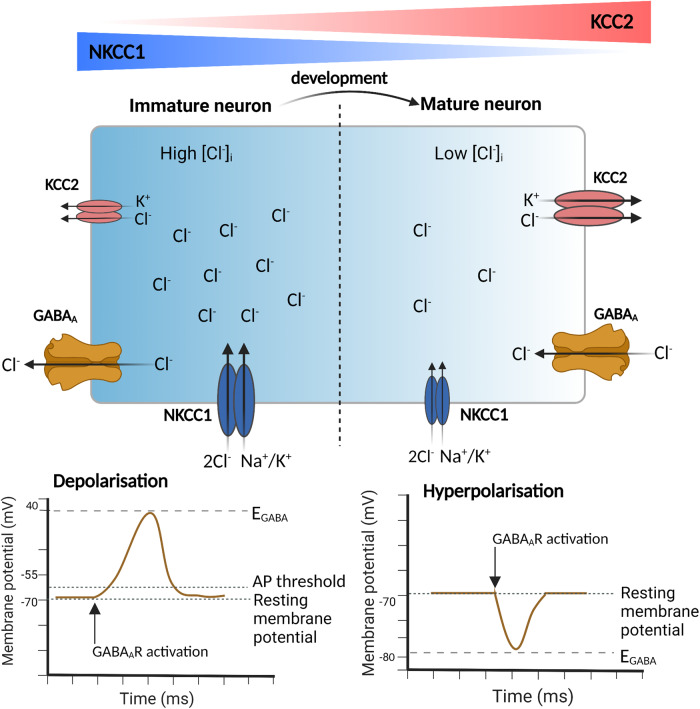
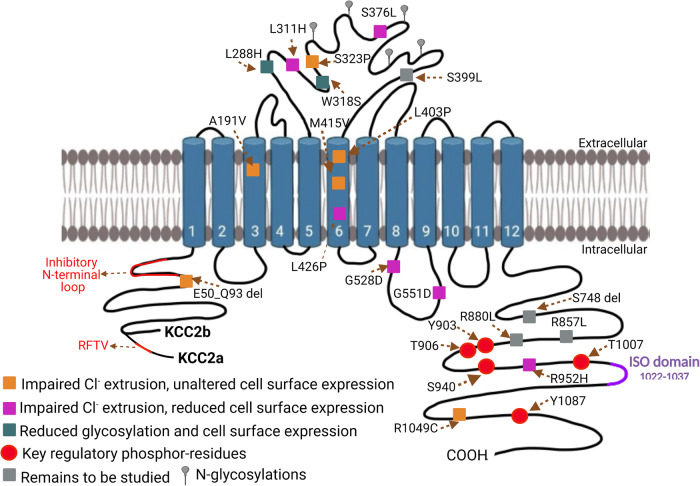
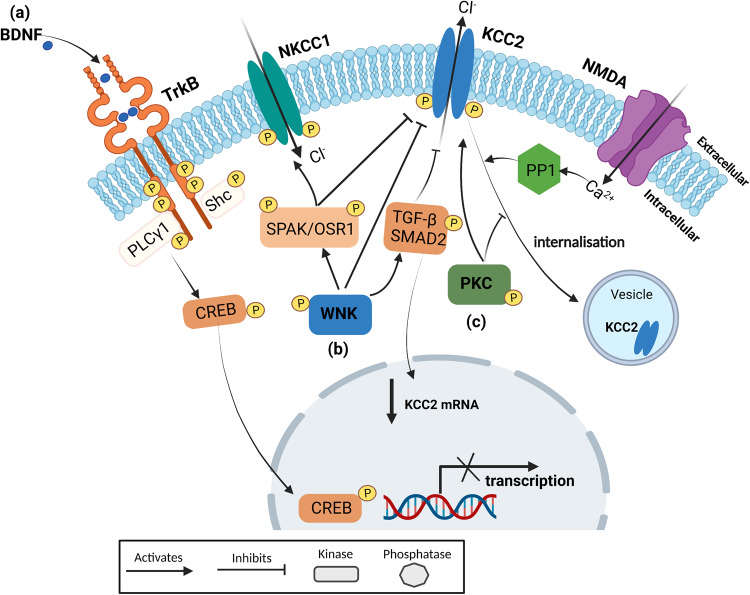
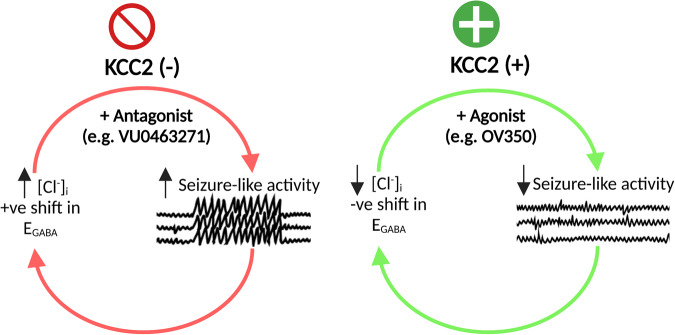
Epilepsy is a prevalent neurological disorder characterized by unprovoked seizures. γ-Aminobutyric acid (GABA) serves as the primary fast inhibitory neurotransmitter in the brain, and GABA binding to the GABAA receptor (GABAAR) regulates Cl- and bicarbonate (HCO3-) influx or efflux through the channel pore, leading to GABAergic inhibition or excitation, respectively. The neuron-specific K+-Cl- cotransporter 2 (KCC2) is essential for maintaining a low intracellular Cl- concentration, ensuring GABAAR-mediated inhibition. Impaired KCC2 function results in GABAergic excitation associated with epileptic activity. Loss-of-function mutations and altered expression of KCC2 lead to elevated [Cl-]i and compromised synaptic inhibition, contributing to epilepsy pathogenesis in human patients. KCC2 antagonism studies demonstrate the necessity of limiting neuronal hyperexcitability within the brain, as reduced KCC2 functioning leads to seizure activity. Strategies focusing on direct (enhancing KCC2 activation) and indirect KCC2 modulation (altering KCC2 phosphorylation and transcription) have proven effective in attenuating seizure severity and exhibiting anti-convulsant properties. These findings highlight KCC2 as a promising therapeutic target for treating epilepsy. Recent advances in understanding KCC2 regulatory mechanisms, particularly via signaling pathways such as WNK, PKC, BDNF, and its receptor TrkB, have led to the discovery of novel small molecules that modulate KCC2. Inhibiting WNK kinase or utilizing newly discovered KCC2 agonists has demonstrated KCC2 activation and seizure attenuation in animal models. This review discusses the role of KCC2 in epilepsy and evaluates its potential as a drug target for epilepsy treatment by exploring various strategies to regulate KCC2 activity.
Keywords: GABAergic inhibition; K+-Cl- cotransporter KCC2; chloride homeostasis; epilepsy; signaling regulatory pathways; small molecular compounds.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures