

A new polymodal gating model of the proton-activated chloride channel
- PMID: 37713449
- PMCID: PMC10529583
- DOI: 10.1371/journal.pbio.3002309
A new polymodal gating model of the proton-activated chloride channel
Abstract
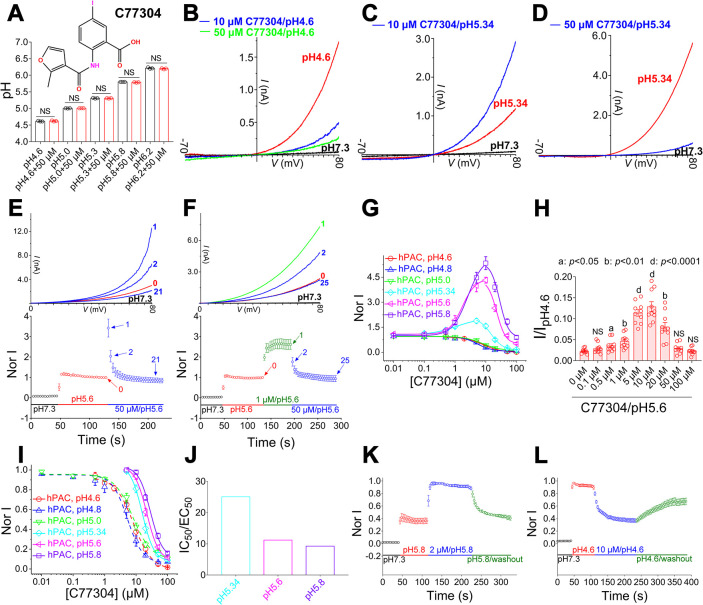
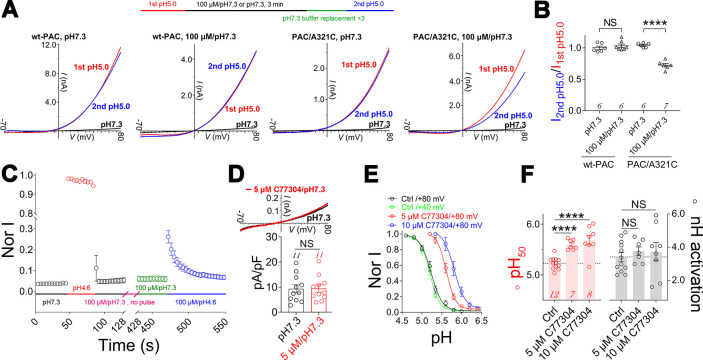
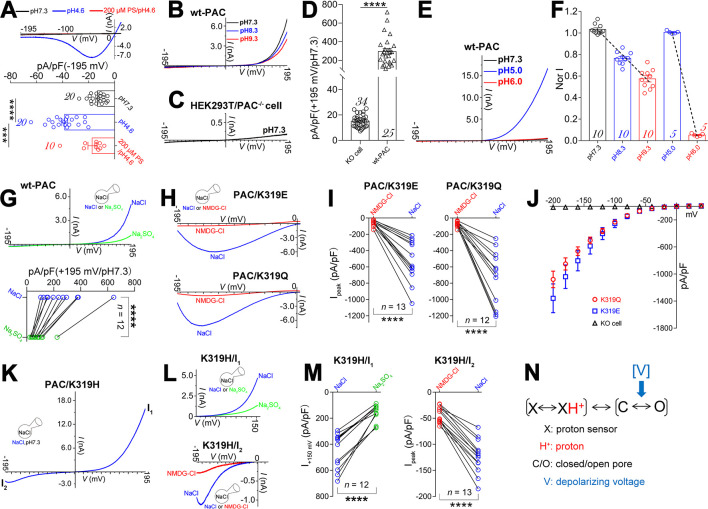
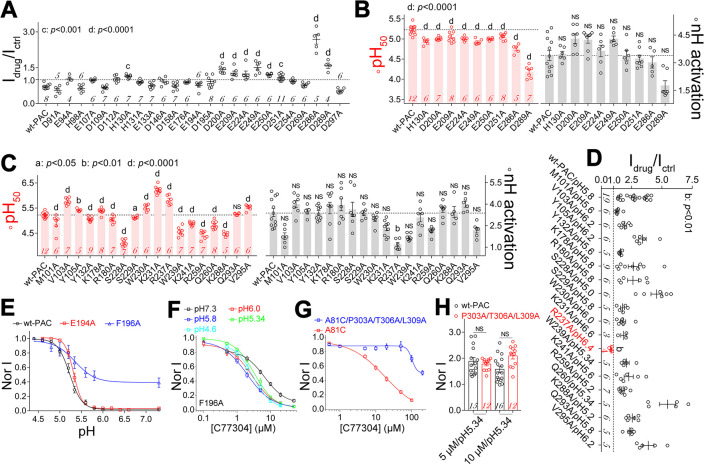
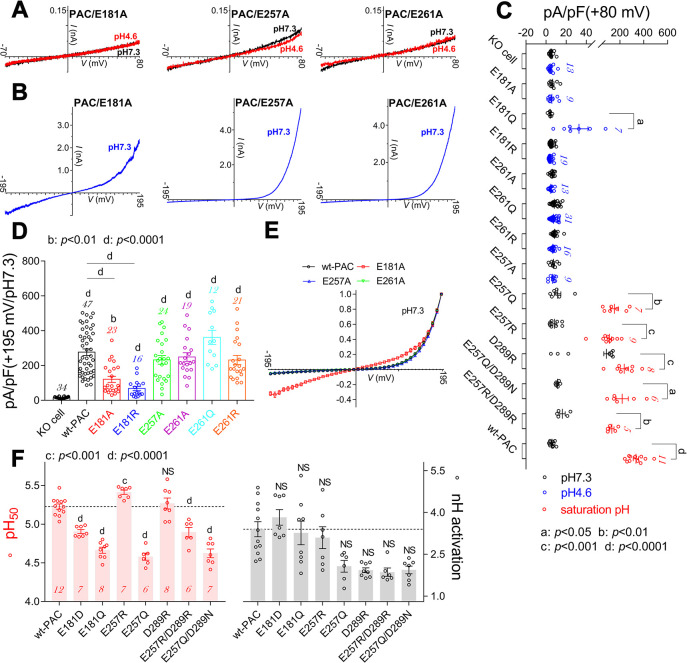
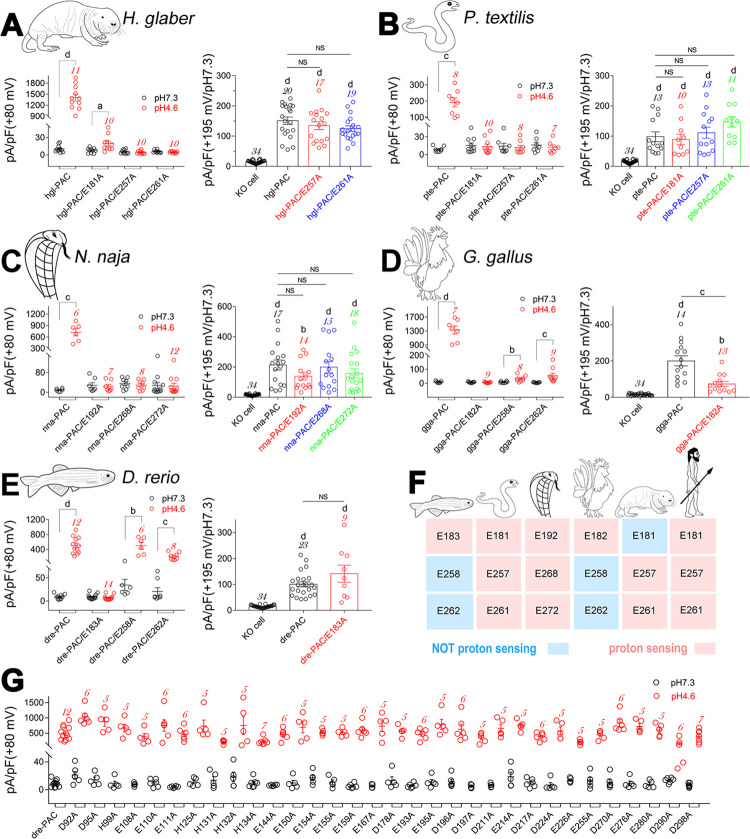
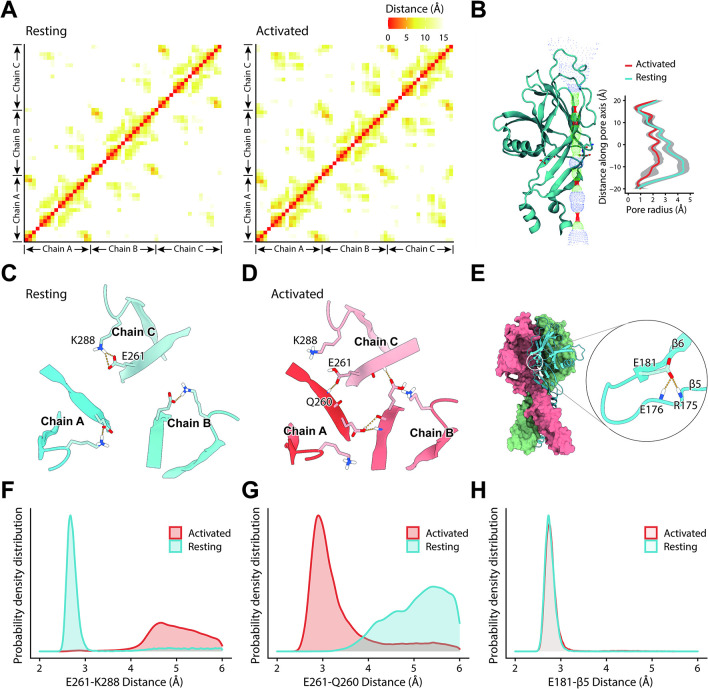
The proton-activated chloride (PAC) channel plays critical roles in ischemic neuron death, but its activation mechanisms remain elusive. Here, we investigated the gating of PAC channels using its novel bifunctional modulator C77304. C77304 acted as a weak activator of the PAC channel, causing moderate activation by acting on its proton gating. However, at higher concentrations, C77304 acted as a weak inhibitor, suppressing channel activity. This dual function was achieved by interacting with 2 modulatory sites of the channel, each with different affinities and dependencies on the channel's state. Moreover, we discovered a protonation-independent voltage activation of the PAC channel that appears to operate through an ion-flux gating mechanism. Through scanning-mutagenesis and molecular dynamics simulation, we confirmed that E181, E257, and E261 in the human PAC channel serve as primary proton sensors, as their alanine mutations eliminated the channel's proton gating while sparing the voltage-dependent gating. This proton-sensing mechanism was conserved among orthologous PAC channels from different species. Collectively, our data unveils the polymodal gating and proton-sensing mechanisms in the PAC channel that may inspire potential drug development.
Copyright: © 2023 Zhao et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
