

Lung tumor-infiltrating Treg have divergent transcriptional profiles and function linked to checkpoint blockade response
- PMID: 37713507
- PMCID: PMC10629528
- DOI: 10.1126/sciimmunol.adg1487
Lung tumor-infiltrating Treg have divergent transcriptional profiles and function linked to checkpoint blockade response
Abstract
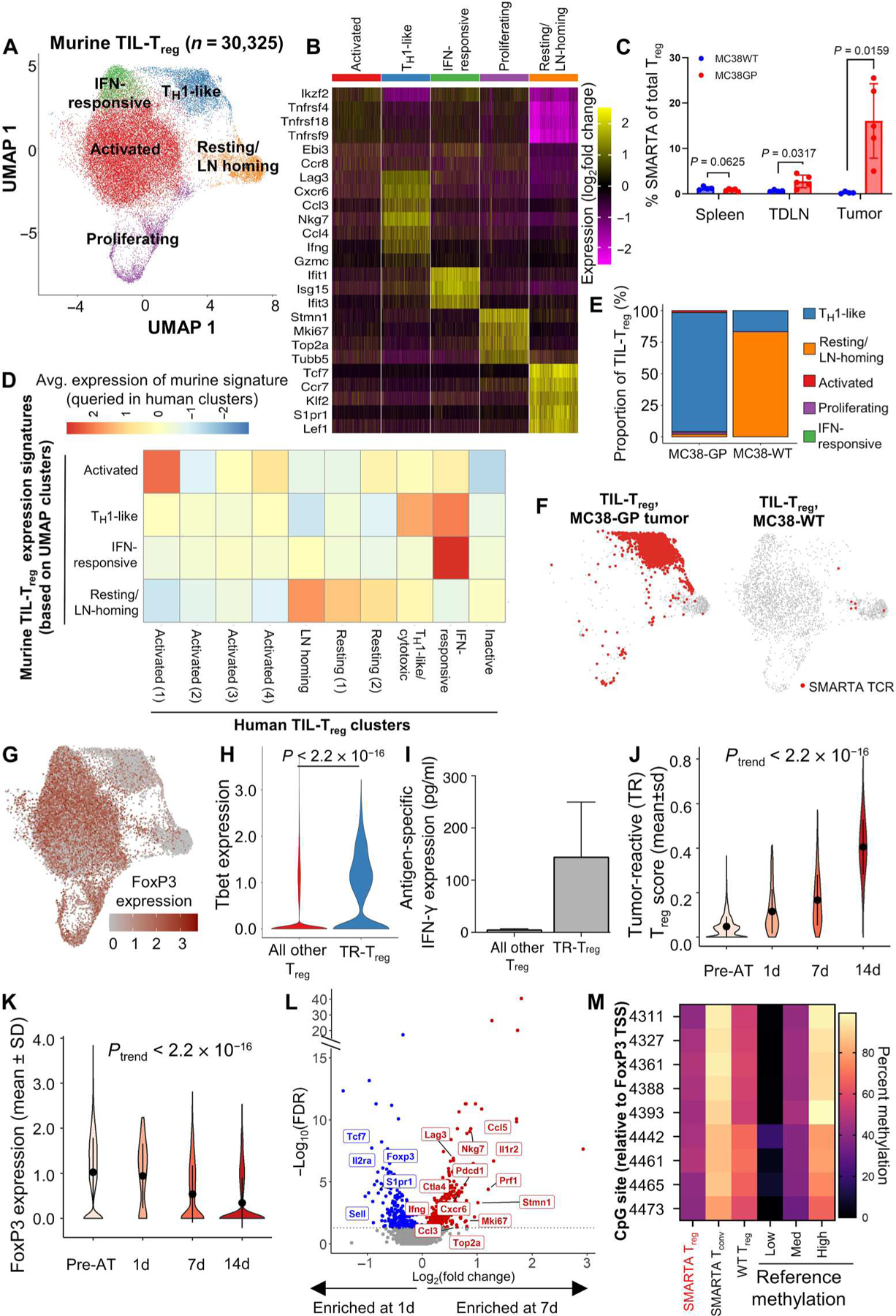
Regulatory T cells (Treg) are conventionally viewed as suppressors of endogenous and therapy-induced antitumor immunity; however, their role in modulating responses to immune checkpoint blockade (ICB) is unclear. In this study, we integrated single-cell RNA-seq/T cell receptor sequencing (TCRseq) of >73,000 tumor-infiltrating Treg (TIL-Treg) from anti-PD-1-treated and treatment-naive non-small cell lung cancers (NSCLC) with single-cell analysis of tumor-associated antigen (TAA)-specific Treg derived from a murine tumor model. We identified 10 subsets of human TIL-Treg, most of which have high concordance with murine TIL-Treg subsets. Only one subset selectively expresses high levels of TNFRSF4 (OX40) and TNFRSF18 (GITR), whose engangement by cognate ligand mediated proliferative programs and NF-κB activation, as well as multiple genes involved in Treg suppression, including LAG3. Functionally, the OX40hiGITRhi subset is the most highly suppressive ex vivo, and its higher representation among total TIL-Treg correlated with resistance to PD-1 blockade. Unexpectedly, in the murine tumor model, we found that virtually all TIL-Treg-expressing T cell receptors that are specific for TAA fully develop a distinct TH1-like signature over a 2-week period after entry into the tumor, down-regulating FoxP3 and up-regulating expression of TBX21 (Tbet), IFNG, and certain proinflammatory granzymes. Transfer learning of a gene score from the murine TAA-specific TH1-like Treg subset to the human single-cell dataset revealed a highly analogous subcluster that was enriched in anti-PD-1-responding tumors. These findings demonstrate that TIL-Treg partition into multiple distinct transcriptionally defined subsets with potentially opposing effects on ICB-induced antitumor immunity and suggest that TAA-specific TIL-Treg may positively contribute to antitumor responses.
Conflict of interest statement
Figures



References
-
- Caushi JX, Zhang J, Ji Z, Vaghasia A, Zhang B, Hsiue EH-C, Mog BJ, Hou W, Justesen S, Blosser R, Tam A, Anagnostou V, Cottrell TR, Guo H, Chan HY, Singh D, Thapa S, Dykema AG, Burman P, Choudhury B, Aparicio L, Cheung LS, Lanis M, Belcaid Z, El Asmar M, Illei PB, Wang R, Meyers J, Schuebel K, Gupta A, Skaist A, Wheelan S, Naidoo J, Marrone KA, Brock M, Ha J, Bush EL, Park BJ, Bott M, Jones DR, Reuss JE, Velculescu VE, Chaft JE, Kinzler KW, Zhou S, Vogelstein B, Taube JM, Hellmann MD, Brahmer JR, Merghoub T, Forde PM, Yegnasubramanian S, Ji H, Pardoll DM, Smith KN, Transcriptional programs of neoantigen-specific TIL in anti-PD-1-treated lung cancers. Nature 596, 126–132 (2021). - PMC - PubMed
-
- Turner JA, Stephen-Victor E, Wang S, Rivas MN, Abdel-Gadir A, Harb H, Cui Y, Fanny M, Charbonnier L-M, Fong JJH, Benamar M, Wang L, Burton OT, Bansal K, Bry L, Zhu C, Li Q-Z, Clement RL, Oettgen HC, Crestani E, Rachid R, Sage PT, Chatila TA, Regulatory T cell-derived TGF-β1 controls multiple checkpoints governing allergy and autoimmunity. Immunity 53, 1202–1214.e6 (2020). - PMC - PubMed
-
- Miyara M, Sakaguchi S, Natural regulatory T cells: Mechanisms of suppression. Trends Mol. Med 13, 108–116 (2007). - PubMed
-
- Nagai H, Horikawa T, Hara I, Fukunaga A, Oniki S, Oka M, Nishigori C, Ichihashi M, In vivo elimination of CD25+ regulatory T cells leads to tumor rejection of B16F10 melanoma, when combined with interleukin-12 gene transfer. Exp. Dermatol 13, 613–620 (2004). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
