

Modular pooled discovery of synthetic knockin sequences to program durable cell therapies
- PMID: 37714135
- PMCID: PMC10508323
- DOI: 10.1016/j.cell.2023.08.013
Modular pooled discovery of synthetic knockin sequences to program durable cell therapies
Abstract
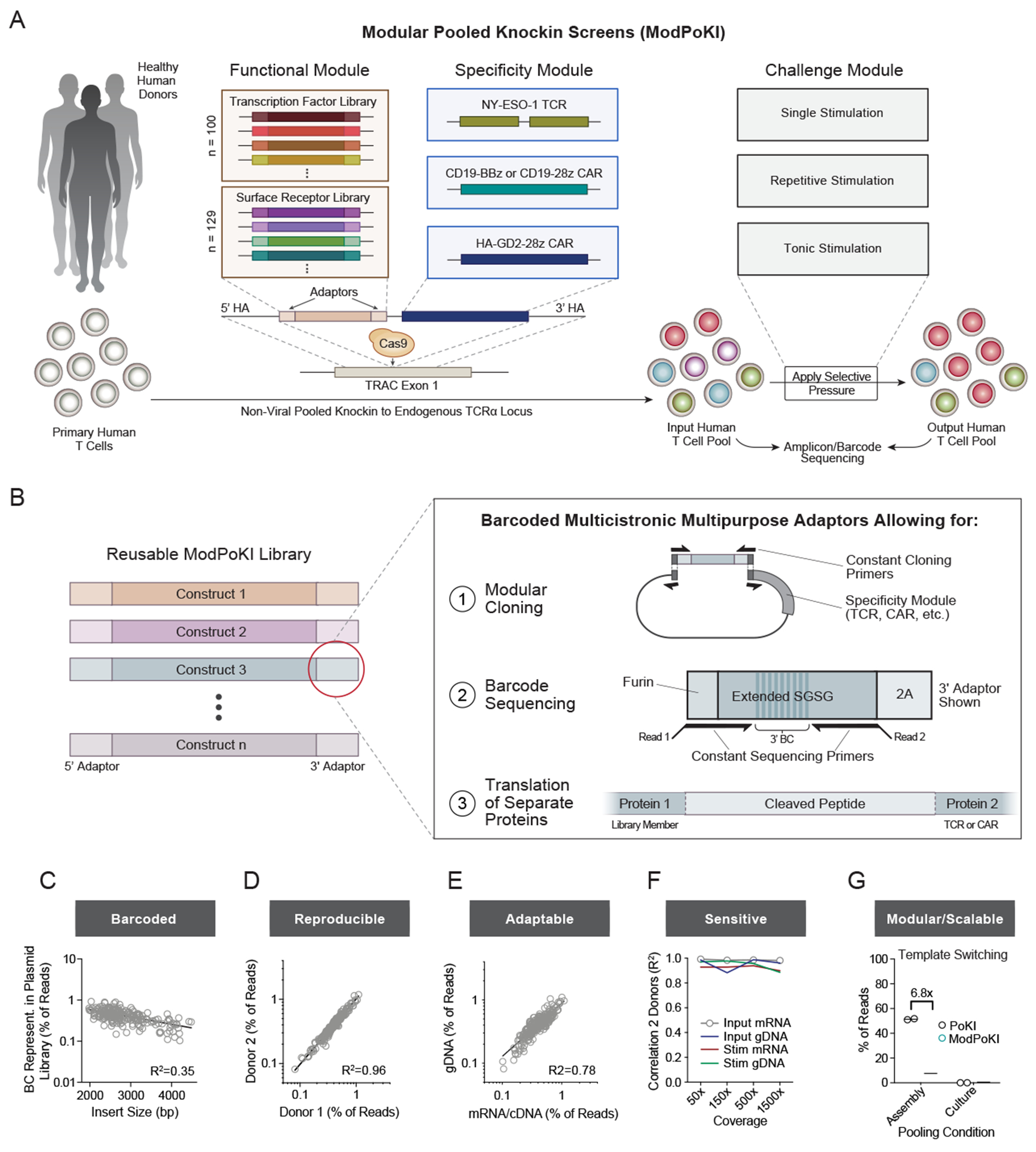
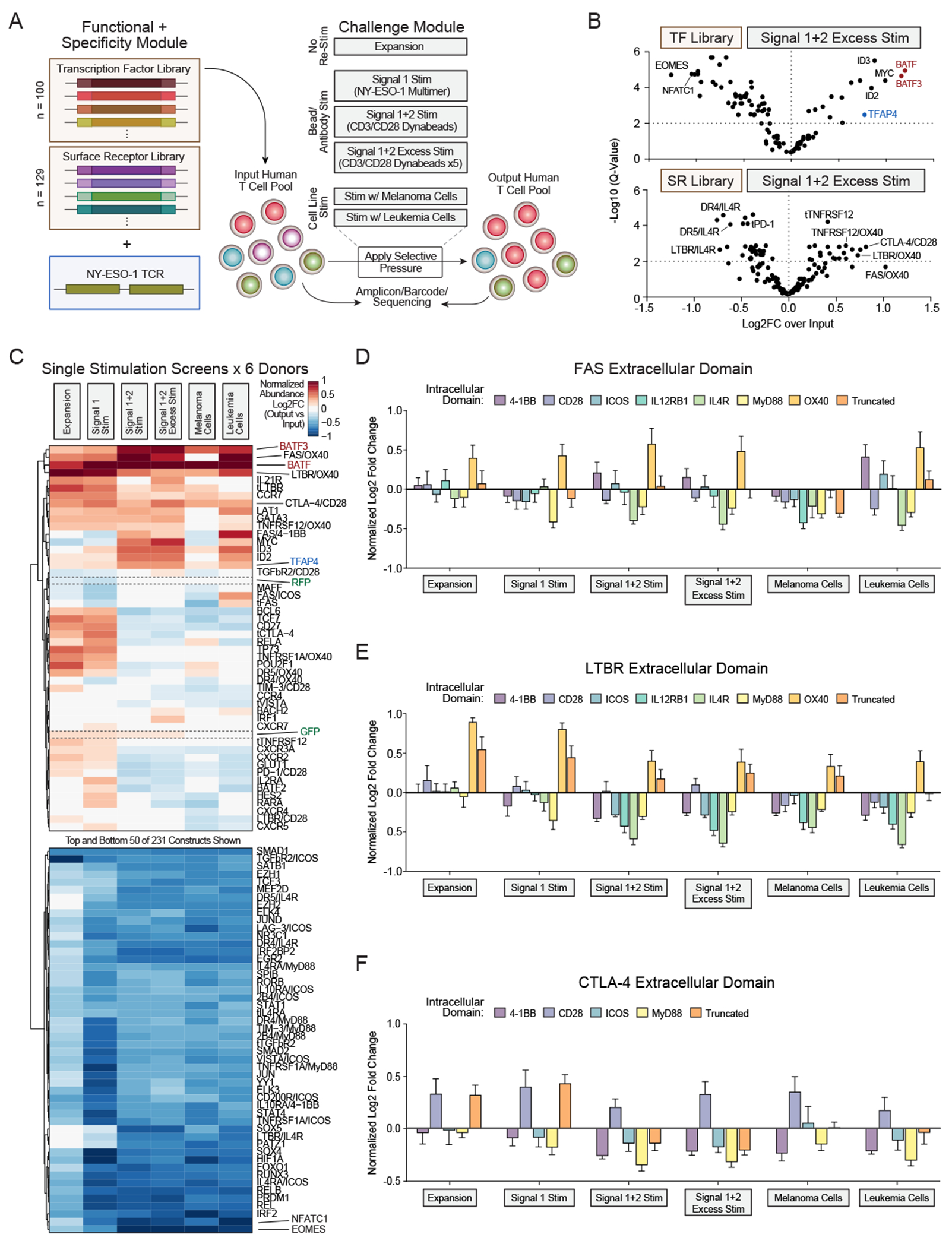
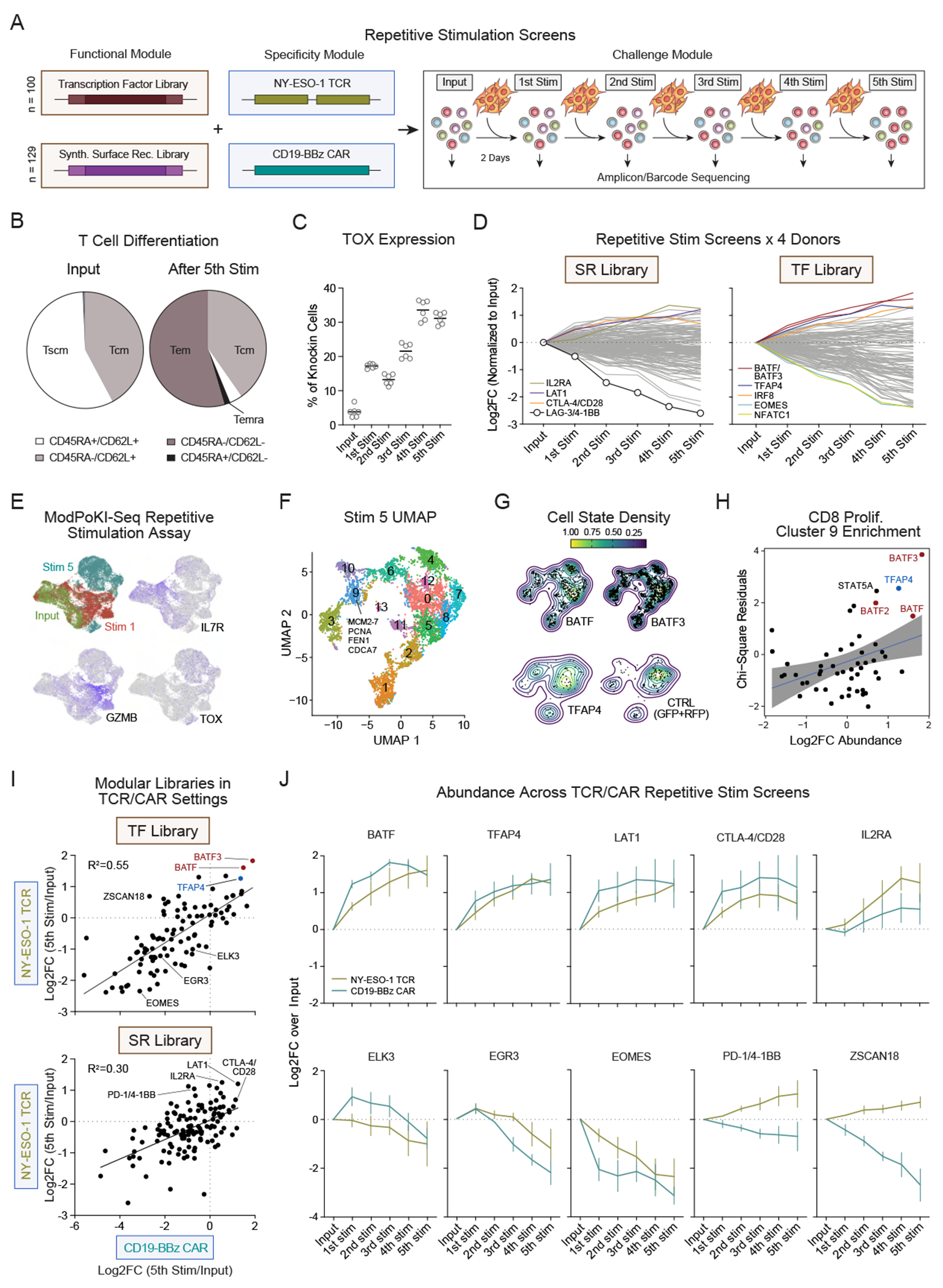
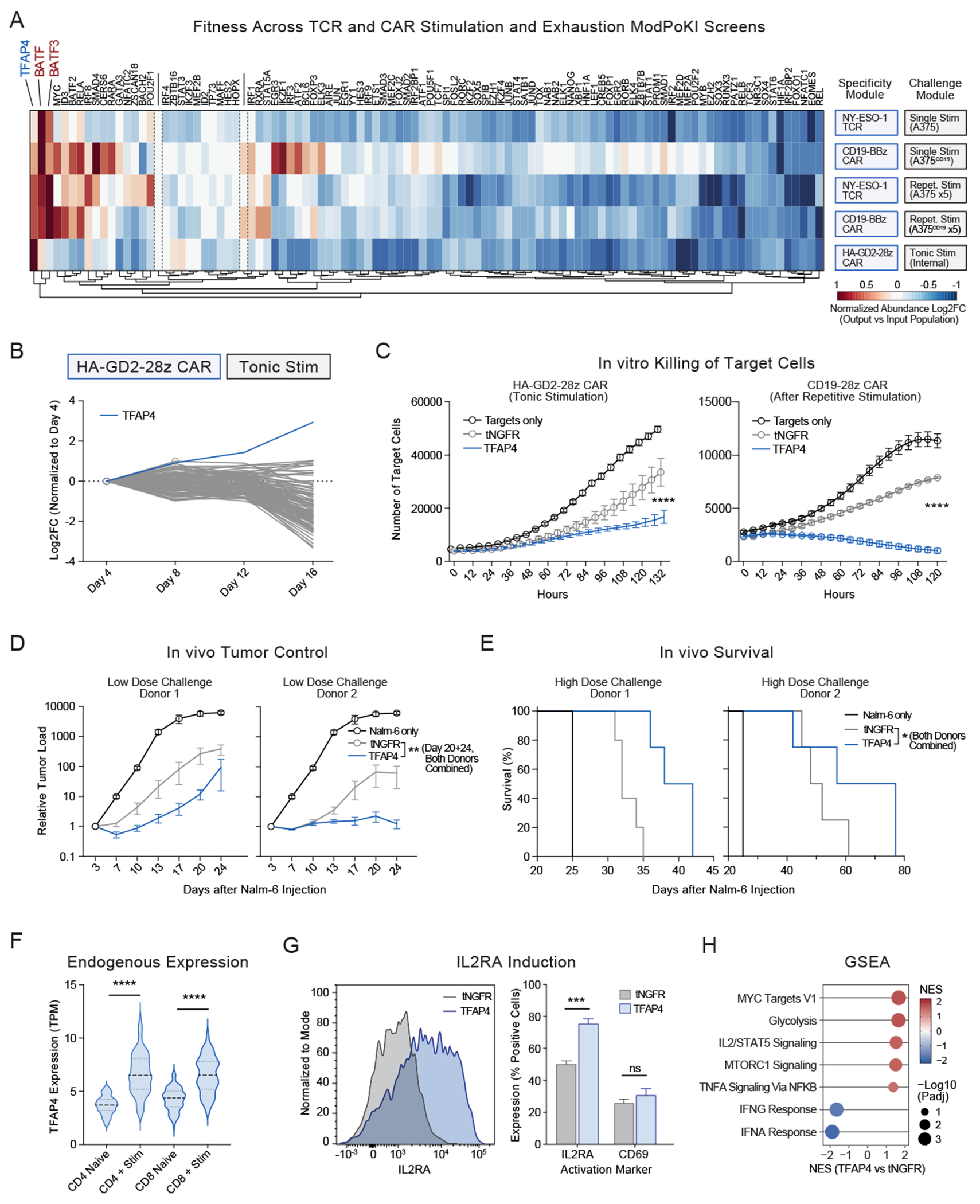
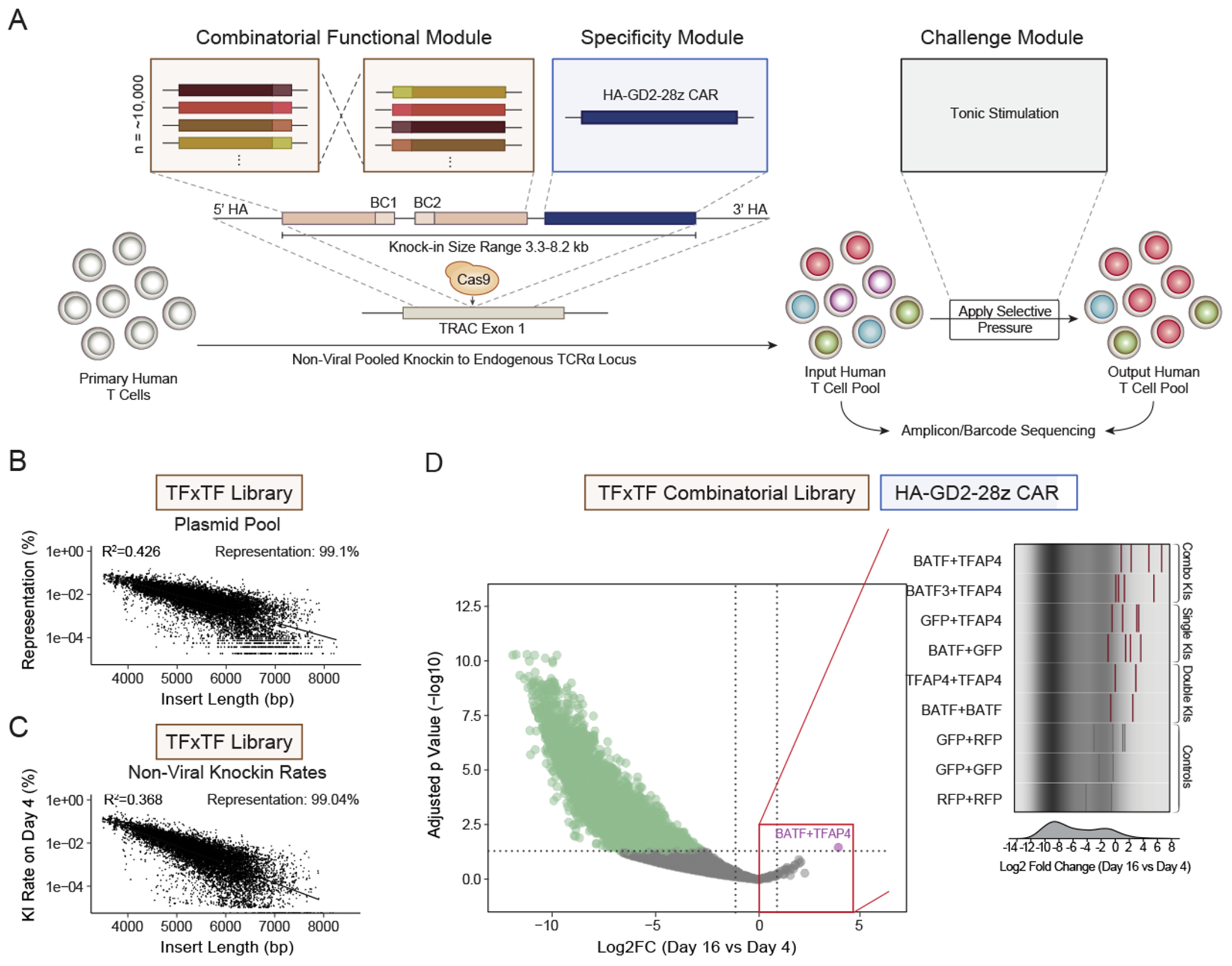
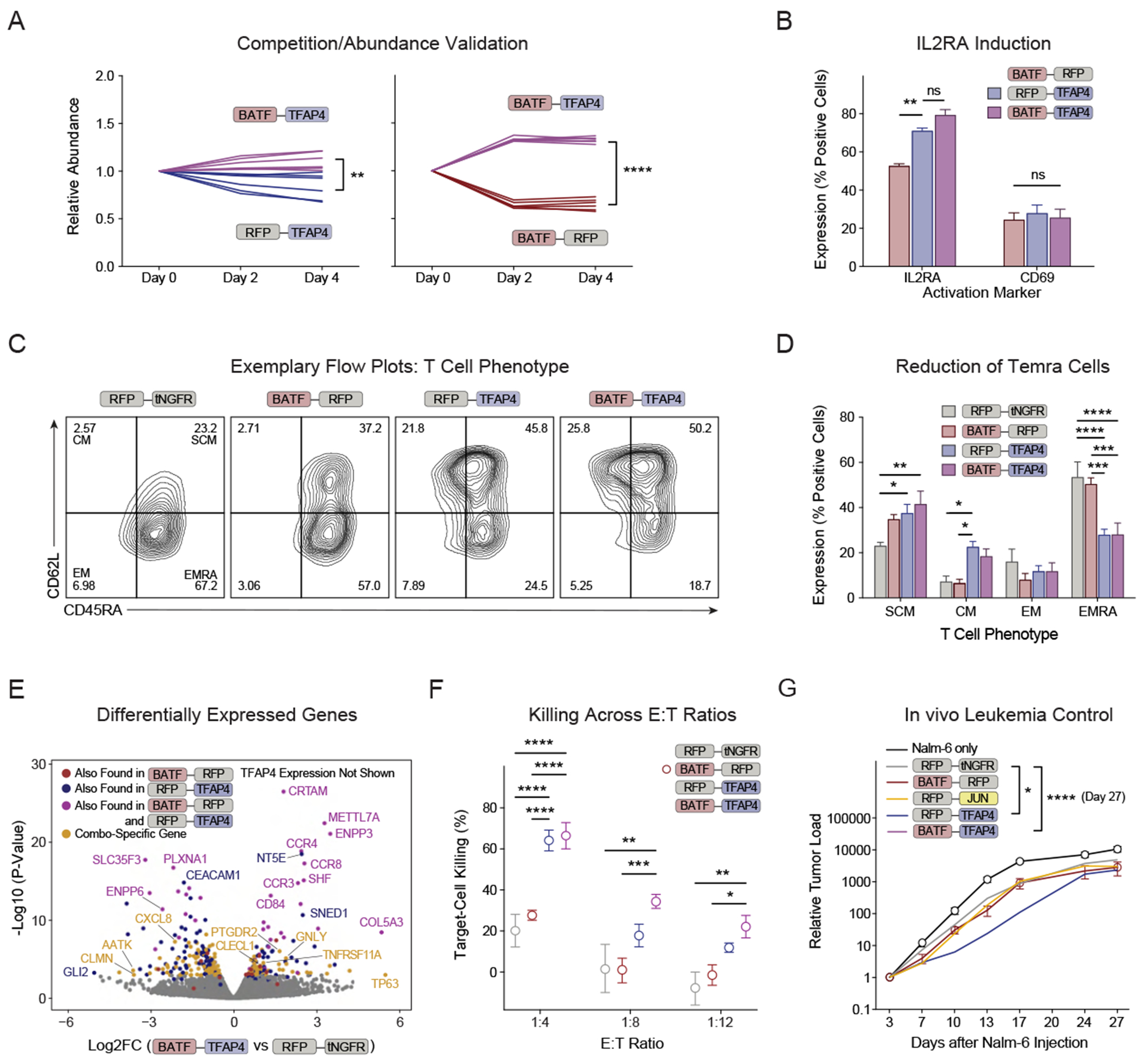
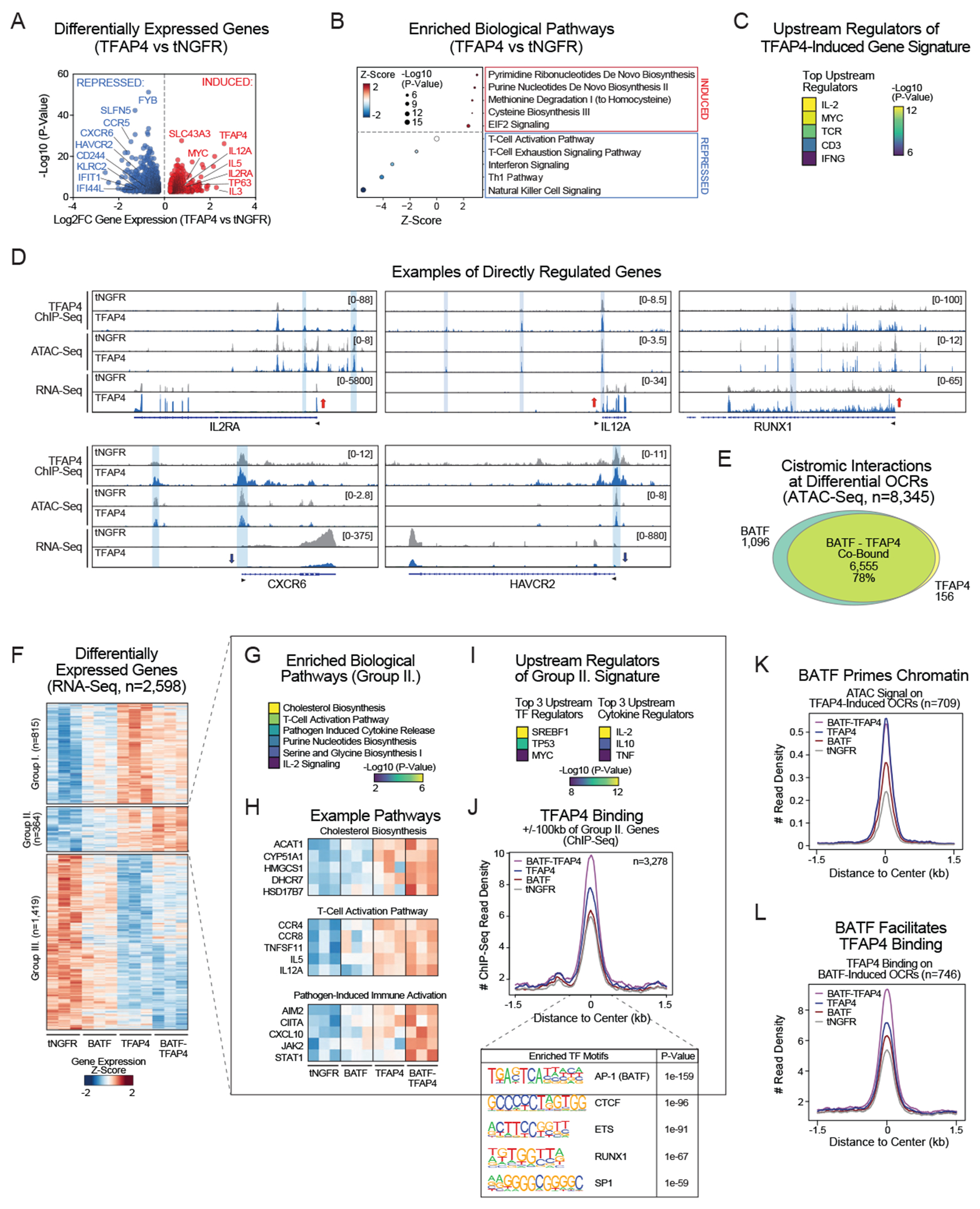
Chronic stimulation can cause T cell dysfunction and limit the efficacy of cellular immunotherapies. Improved methods are required to compare large numbers of synthetic knockin (KI) sequences to reprogram cell functions. Here, we developed modular pooled KI screening (ModPoKI), an adaptable platform for modular construction of DNA KI libraries using barcoded multicistronic adaptors. We built two ModPoKI libraries of 100 transcription factors (TFs) and 129 natural and synthetic surface receptors (SRs). Over 30 ModPoKI screens across human TCR- and CAR-T cells in diverse conditions identified a transcription factor AP4 (TFAP4) construct that enhanced fitness of chronically stimulated CAR-T cells and anti-cancer function in vitro and in vivo. ModPoKI's modularity allowed us to generate an ∼10,000-member library of TF combinations. Non-viral KI of a combined BATF-TFAP4 polycistronic construct enhanced fitness. Overexpressed BATF and TFAP4 co-occupy and regulate key gene targets to reprogram T cell function. ModPoKI facilitates the discovery of complex gene constructs to program cellular functions.
Keywords: CRISPR; chimeric antigen receptor; chronic stimulation; human T cells; immunotherapy; knockins; pooled screens; synthetic surface receptor; transcription factor.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests F.B. received research awards (Gilead and Kite and Bristol Myers Squibb Foundation Immunonkologie). E.S. was an advisor for Arsenal Biosciences. J.E. is a compensated co-founder at Mnemo Therapeutics and compensated scientific advisor to Cytovia Therapeutics. J.E. owns stocks in Mnemo Therapeutics and Cytovia Therapeutics. J.E. has received a consulting fee from Casdin Capital. The Eyquem lab has received research support from Cytovia Therapeutics and Takeda. T.L.R. is a compensated co-founder, member of the scientific advisory board, and previously worked as the CSO of Arsenal Biosciences. A.T.S. is a founder of Immunai and Cartography Biosciences and receives research funding from Allogene Therapeutics and Merck Research Laboratories. C.T.M. is a compensated Bio+Health Venture Fellow at Andreessen Horowitz. C.J.Y. is founder for and holds equity in DropPrint Genomics (now ImmunAI) and Survey Genomics, a scientific advisory board member for and holds equity in Related Sciences and ImmunAI, a consultant for and holds equity in Maze Therapeutics, and a consultant for TReX Bio, HiBio, ImYoo, and Santa Ana. C.J.Y. has received research support from Chan Zuckerberg Initiative, Chan Zuckerberg Biohub, Genentech, BioLegend, ScaleBio, and Illumina. A.M. is a co-founder of Arsenal Biosciences, Spotlight Therapeutics, and Survey Genomics, serves on the boards of directors at Spotlight Therapeutics and Survey Genomics, is a board observer (and former member of the board of directors) at Arsenal Biosciences, is a member of the scientific advisory boards of Arsenal Biosciences, Spotlight Therapeutics, Survey Genomics, NewLimit, Amgen, Tenaya, and Lightcast, owns stock in Arsenal Biosciences, Spotlight Therapeutics, NewLimit, Survey Genomics, PACT Pharma, Tenaya, and Lightcast and has received fees from Arsenal Biosciences, Spotlight Therapeutics, Survey Genomics, NewLimit, 23andMe, PACT Pharma, Juno Therapeutics, Tenaya, Lightcast, GLG, Gilead, Trizell, Vertex, Merck, Amgen, Genentech, AlphaSights, Rupert Case Management, Bernstein, and ALDA. A.M. is an investor in and informal advisor to Offline Ventures and a client of EPIQ. The Marson laboratory received research support from Juno Therapeutics, Epinomics, Sanofi, GlaxoSmithKline, Gilead, and Anthem. T.L.R., F.B., A.M., R.A., Y.Y.C., C.T.M., and E.S. are listed on patent applications related to this work.
Figures
References
Publication types
MeSH terms
Grants and funding
- F30 DK120213/DK/NIDDK NIH HHS/United States
- P30 DK063720/DK/NIDDK NIH HHS/United States
- T32 GM007618/GM/NIGMS NIH HHS/United States
- S10 RR028962/RR/NCRR NIH HHS/United States
- K08 CA286740/CA/NCI NIH HHS/United States
- T32 CA108462/CA/NCI NIH HHS/United States
- K08 CA252605/CA/NCI NIH HHS/United States
- R01 HG011239/HG/NHGRI NIH HHS/United States
- T32 AI007334/AI/NIAID NIH HHS/United States
- T32 DK007418/DK/NIDDK NIH HHS/United States
- S10 OD028511/OD/NIH HHS/United States
- R01 CA276368/CA/NCI NIH HHS/United States
- S10 OD021822/OD/NIH HHS/United States
- R01 AI136972/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
