

A linear and circular dual-conformation noncoding RNA involved in oxidative stress tolerance in Bacillus altitudinis
- PMID: 37714854
- PMCID: PMC10504365
- DOI: 10.1038/s41467-023-41491-4
A linear and circular dual-conformation noncoding RNA involved in oxidative stress tolerance in Bacillus altitudinis
Abstract
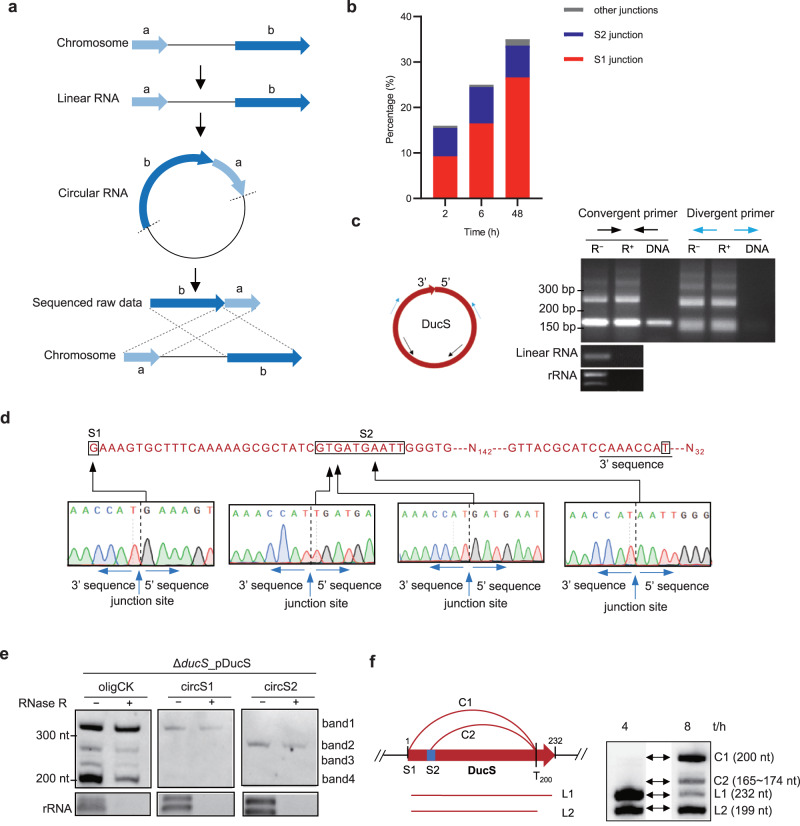
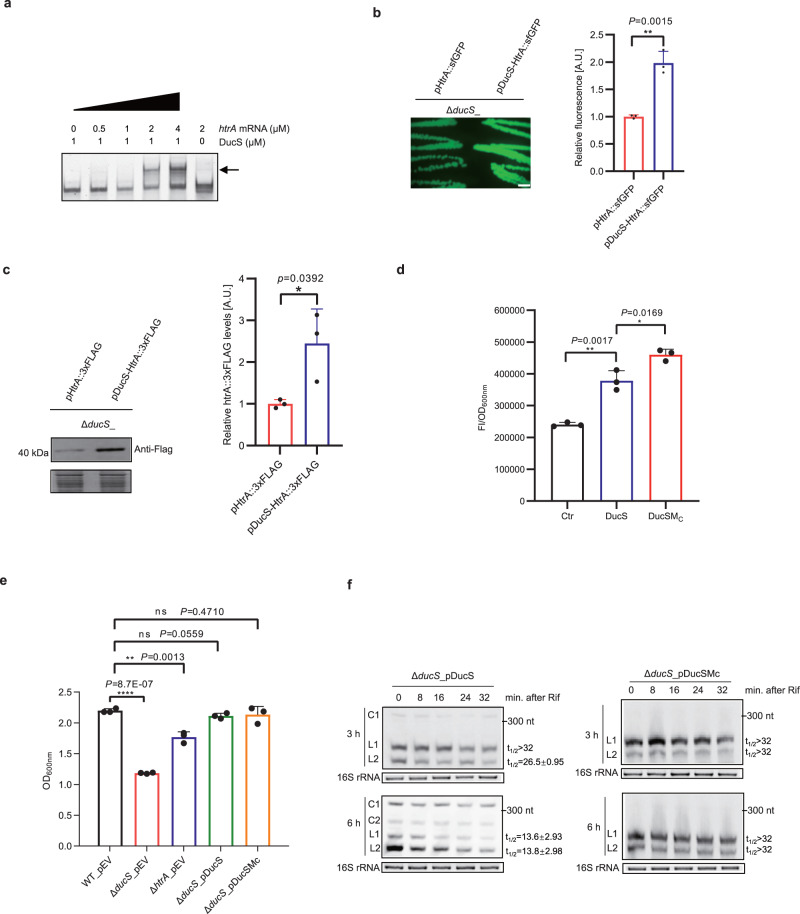
Circular RNAs have been extensively studied in eukaryotes, but their presence and/or biological functionality in bacteria are unclear. Here, we show that a regulatory noncoding RNA (DucS) exists in both linear and circular conformation in Bacillus altitudinis. The linear forms promote B. altitudinis tolerance to H2O2 stress, partly through increased translation of a stress-responsive gene, htrA. The 3' end sequences of the linear forms are crucial for RNA circularization, and formation of circular forms can decrease the levels of the regulatory linear cognates. Bioinformatic analysis of available RNA-seq datasets from 30 bacterial species revealed multiple circular RNA candidates, distinct from DucS, for all the examined species. Experiments testing for the presence of selected circular RNA candidates in four species successfully validated 7 out of 9 candidates from B. altitudinis and 4 out of 5 candidates from Bacillus paralicheniformis; However, none of the candidates tested for Bacillus subtilis and Escherichia coli were detected. Our work identifies a dual-conformation regulatory RNA in B. altitutidinis, and indicates that circular RNAs exist in diverse bacteria. However, circularization of specific RNAs does not seem to be conserved across species, and the circularization mechanisms and biological functionality of the circular forms remain unclear.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
CircANKRD12 Is Induced in Endothelial Cell Response to Oxidative Stress.Cells. 2022 Nov 9;11(22):3546. doi: 10.3390/cells11223546. Cells. 2022. PMID: 36428974 Free PMC article.
-
Circular ribozymes generated in Escherichia coli using group I self-splicing permuted intron-exon sequences.J Biol Chem. 1996 Oct 18;271(42):26081-7. doi: 10.1074/jbc.271.42.26081. J Biol Chem. 1996. PMID: 8824250
-
6S RNA, a global regulator of transcription in Escherichia coli, Bacillus subtilis, and beyond.Annu Rev Microbiol. 2014;68:45-60. doi: 10.1146/annurev-micro-092611-150135. Epub 2014 Apr 10. Annu Rev Microbiol. 2014. PMID: 24742053 Review.
-
Endophytic Bacillus altitudinis WR10 alleviates Cu toxicity in wheat by augmenting reactive oxygen species scavenging and phenylpropanoid biosynthesis.J Hazard Mater. 2021 Mar 5;405:124272. doi: 10.1016/j.jhazmat.2020.124272. Epub 2020 Oct 13. J Hazard Mater. 2021. PMID: 33097348
-
Emerging Clues of Regulatory Roles of Circular RNAs through Modulating Oxidative Stress: Focus on Neurological and Vascular Diseases.Oxid Med Cell Longev. 2021 Mar 1;2021:6659908. doi: 10.1155/2021/6659908. eCollection 2021. Oxid Med Cell Longev. 2021. PMID: 33747348 Free PMC article. Review.
Cited by
-
Circular RNA vaccines: Pioneering the next-gen cancer immunotherapy.Cancer Pathog Ther. 2024 Dec 4;3(4):309-321. doi: 10.1016/j.cpt.2024.11.003. eCollection 2025 Jul. Cancer Pathog Ther. 2024. PMID: 40697518 Free PMC article. Review.
-
Species-specific circular RNA circDS-1 enhances adaptive evolution in Talaromyces marneffei through regulation of dimorphic transition.PLoS Genet. 2025 Mar 6;21(3):e1011482. doi: 10.1371/journal.pgen.1011482. eCollection 2025 Mar. PLoS Genet. 2025. PMID: 40048447 Free PMC article.
-
Circular RNAs in Archaea.Adv Exp Med Biol. 2025;1485:451-464. doi: 10.1007/978-981-96-9428-0_26. Adv Exp Med Biol. 2025. PMID: 40886291 Review.
-
Emerging Roles of Circular RNA in Macrophage Activation and Inflammatory Lung Responses.Cells. 2024 Aug 23;13(17):1407. doi: 10.3390/cells13171407. Cells. 2024. PMID: 39272979 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
