

Metabolic modulation of mitochondrial mass during CD4+ T cell activation
- PMID: 37716347
- PMCID: PMC10604707
- DOI: 10.1016/j.chembiol.2023.08.008
Metabolic modulation of mitochondrial mass during CD4+ T cell activation
Abstract
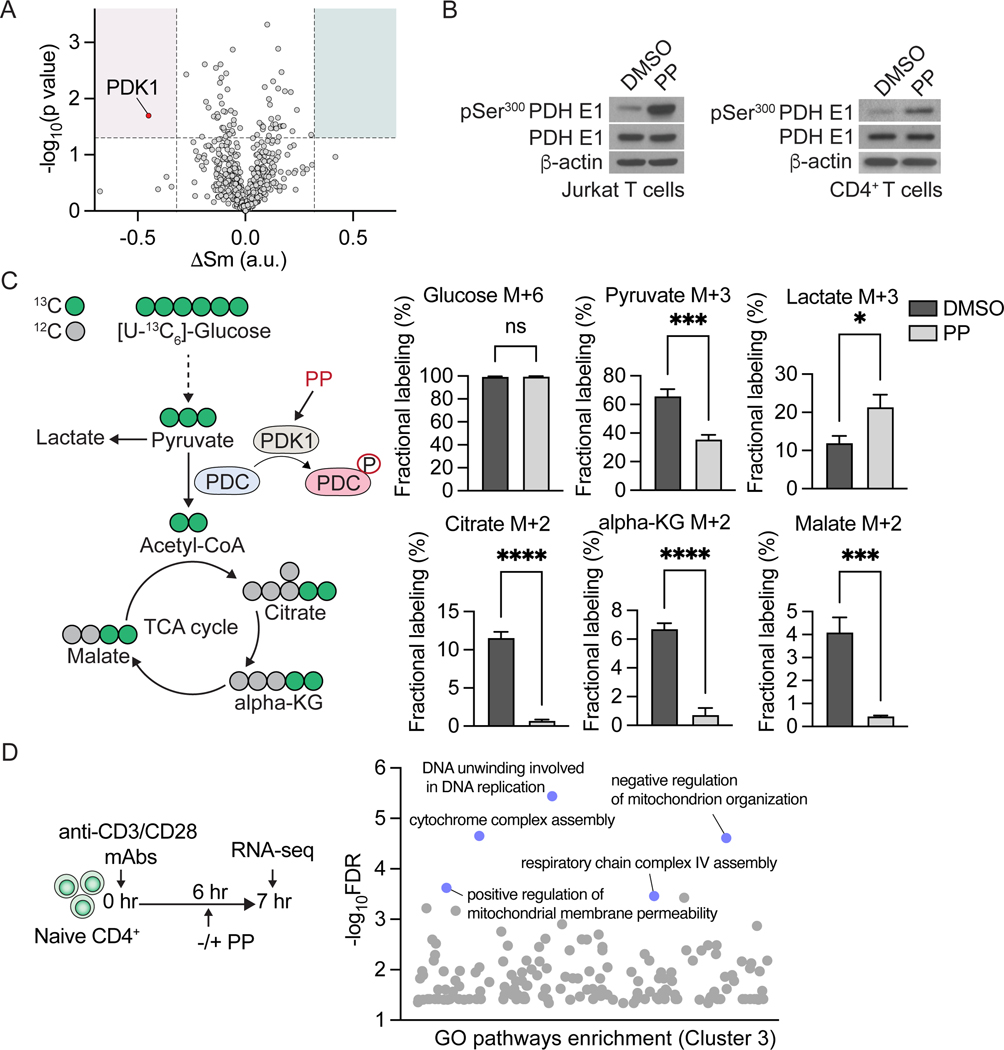
Mitochondrial biogenesis initiates within hours of T cell receptor (TCR) engagement and is critical for T cell activation, function, and survival; yet, how metabolic programs support mitochondrial biogenesis during TCR signaling is not fully understood. Here, we performed a multiplexed metabolic chemical screen in CD4+ T lymphocytes to identify modulators of metabolism that impact mitochondrial mass during early T cell activation. Treatment of T cells with pyrvinium pamoate early during their activation blocks an increase in mitochondrial mass and results in reduced proliferation, skewed CD4+ T cell differentiation, and reduced cytokine production. Furthermore, administration of pyrvinium pamoate at the time of induction of experimental autoimmune encephalomyelitis, an experimental model of multiple sclerosis in mice, prevented the onset of clinical disease. Thus, modulation of mitochondrial biogenesis may provide a therapeutic strategy for modulating T cell immune responses.
Keywords: CD4(+) T cells; T cell differentiation; high-throughput metabolic screen; mitochondrial biogenesis; pyruvate oxidation; pyrvinium pamoate.
Copyright © 2023 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of interests M.C.H. serves on the advisory boards for Alixia, Minovia, and MitoQ. M.C.H. has received funding from Agilent and Roche in the last five years. M.C.H. and K.K. have patents pending on the role of PP in T cells and mitochondria. A.S. is an employee of Flagship Labs 84, Inc, a subsidiary of Flagship Pioneering. J.M.D. reports grants from NIH during the conduct of the study, as well as personal fees from ElevateBio (consulting) and Third Rock Ventures (consulting) outside the submitted work. S.H. has consulted for Merck KGaA. I.S.H. reports financial support from Kojin Therapeutics and consulting fees for Ono Pharma USA. Fundings and fees from these companies are outside the scope of the current work. P.G. reports personal fees from RA Capital for consulting outside the submitted work. P.K.S. is a member of the SAB or BOD member of Applied Biomath, RareCyte Inc., and Glencoe Software; P.K.S. is also a member of the NanoString SAB, and the Sorger laboratory has received research funding from Novartis and Merck in the last five years. P.K.S. declares that none of these relationships have influenced the content of this manuscript. S.K.D. receives research funding unrelated to this project from Novartis and BMS and is a founder and scientific advisory board member for Kojin. A.H.S. has funding from Quark, Merck, AbbVie, Moderna, and Vertex unrelated to the submitted work. A.H.S. serves on advisory boards for SQZ Biotechnologies, Selecta, Elpiscience, Monopteros, Bicara, Fibrogen, Alixia, IOME, Corner Therapeutics, Glaxo Smith Kline, Amgen, and Janssen. She also is on scientific advisory boards for the Massachusetts General Cancer Center, Program in Cellular and Molecular Medicine at Boston Children’s Hospital, the Human Oncology and Pathogenesis Program at Memorial Sloan Kettering Cancer Center, the Gladstone Institute, and the Johns Hopkins Bloomberg-Kimmel Institute for Cancer Immunotherapy. She is an academic editor for the Journal of Experimental Medicine. A.H.S. has patents/pending royalties on the PD-1 pathway from Roche and Novartis.
Figures




References
-
- Vardhana SA, Hwee MA, Berisa M, Wells DK, Yost KE, King B, Smith M, Herrera PS, Chang HY, Satpathy AT, et al. (2020). Impaired mitochondrial oxidative phosphorylation limits the self-renewal of T cells exposed to persistent antigen. Nat Immunol 21, 1022–1033. 10.1038/s41590-020-0725-2. - DOI - PMC - PubMed
-
- Akkaya B, Roesler AS, Miozzo P, Theall BP, Al Souz J, Smelkinson MG, Kabat J, Traba J, Sack MN, Brzostowski JA, et al. (2018). Increased Mitochondrial Biogenesis and Reactive Oxygen Species Production Accompany Prolonged CD4(+) T Cell Activation. J Immunol 201, 3294–3306. 10.4049/jimmunol.1800753. - DOI - PMC - PubMed
-
- Ron-Harel N, Santos D, Ghergurovich JM, Sage PT, Reddy A, Lovitch SB, Dephoure N, Satterstrom FK, Sheffer M, Spinelli JB, et al. (2016). Mitochondrial Biogenesis and Proteome Remodeling Promote One-Carbon Metabolism for T Cell Activation. Cell Metab 24, 104–117. 10.1016/j.cmet.2016.06.007. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
