

Hypoxia-inducible factor upregulation by roxadustat attenuates drug reward by altering brain iron homoeostasis
- PMID: 37718358
- PMCID: PMC10505610
- DOI: 10.1038/s41392-023-01578-2
Hypoxia-inducible factor upregulation by roxadustat attenuates drug reward by altering brain iron homoeostasis
Abstract
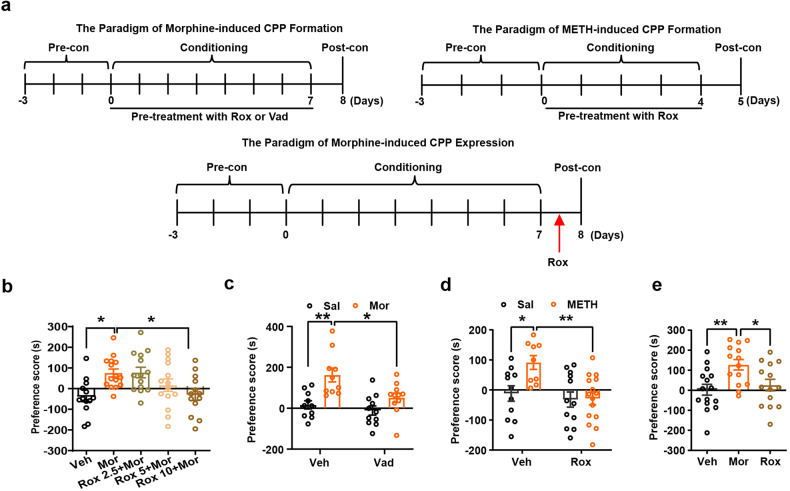
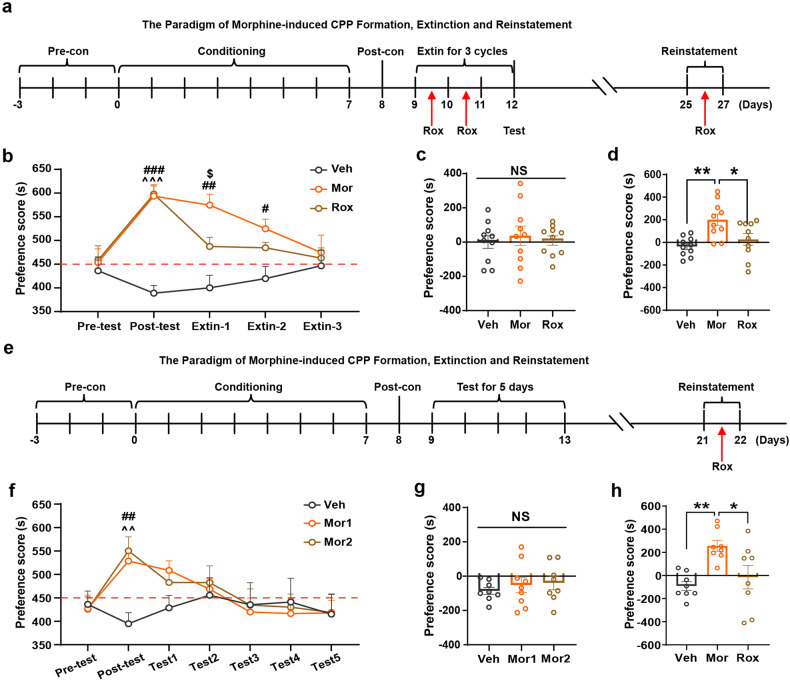
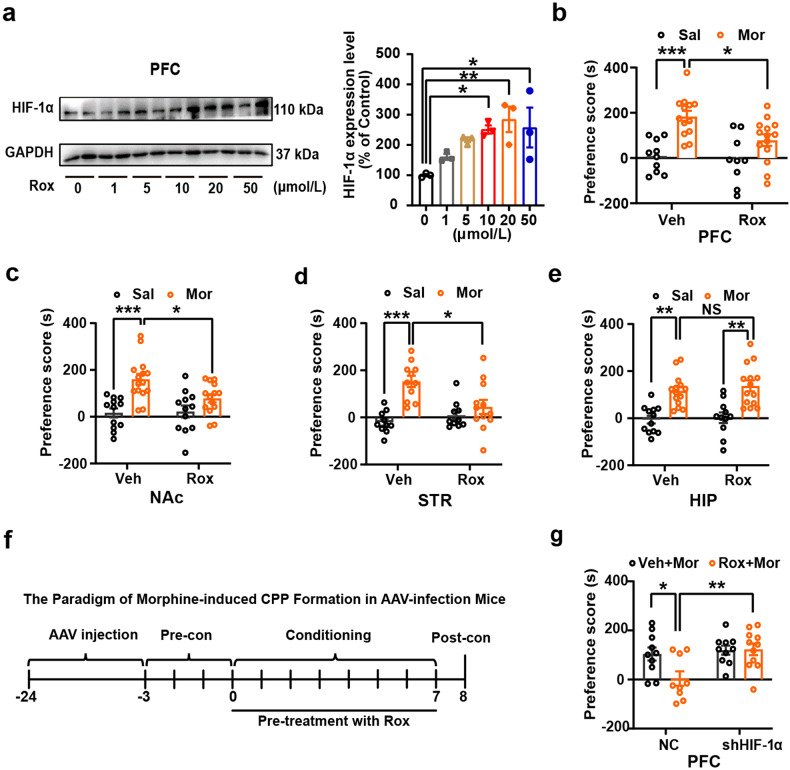
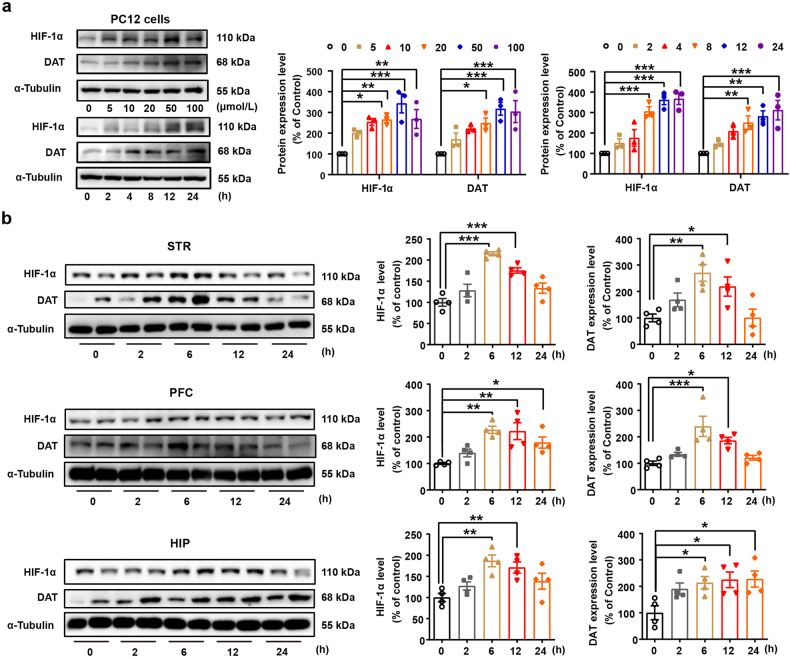
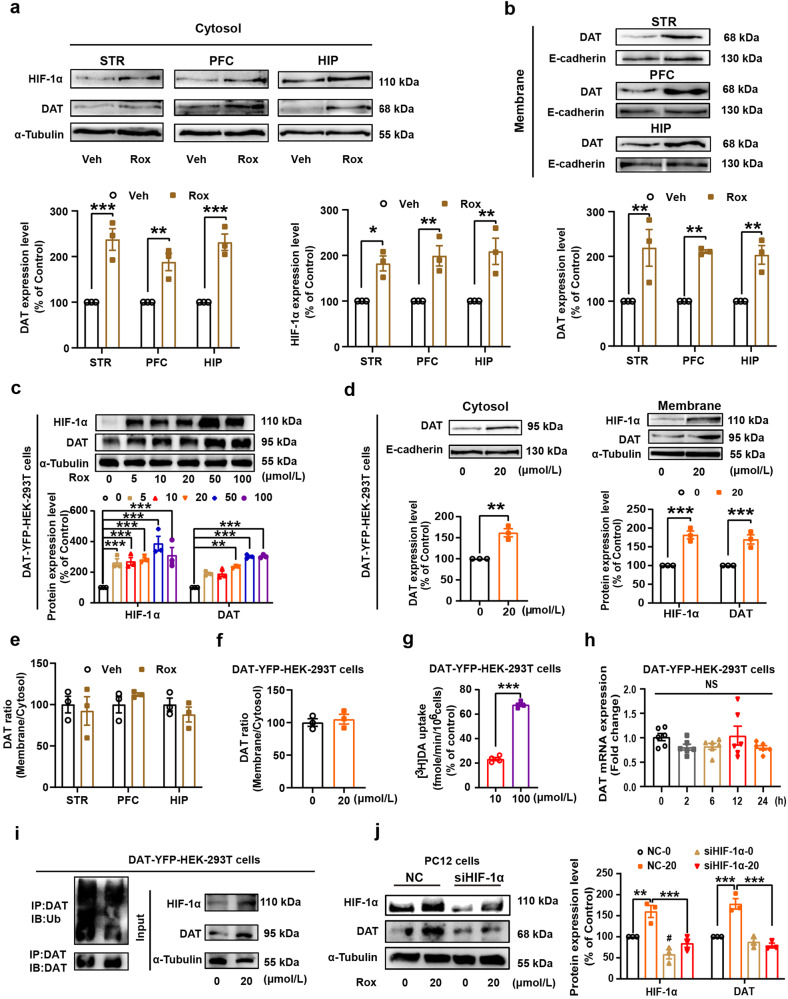
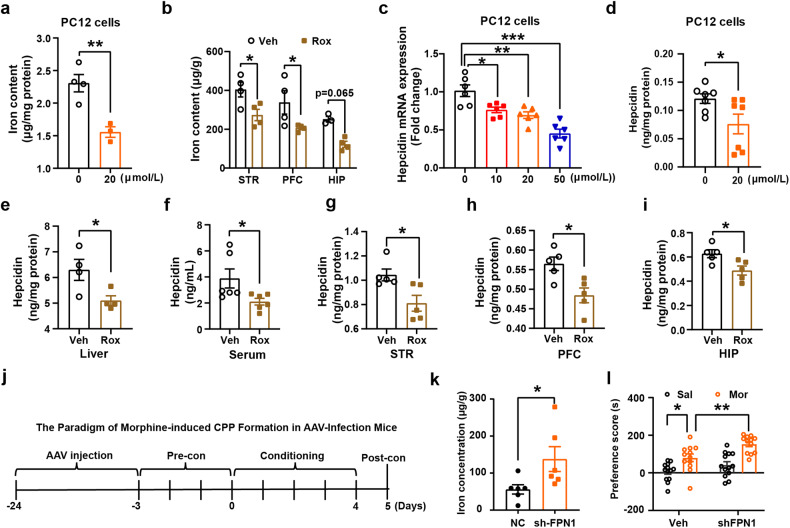
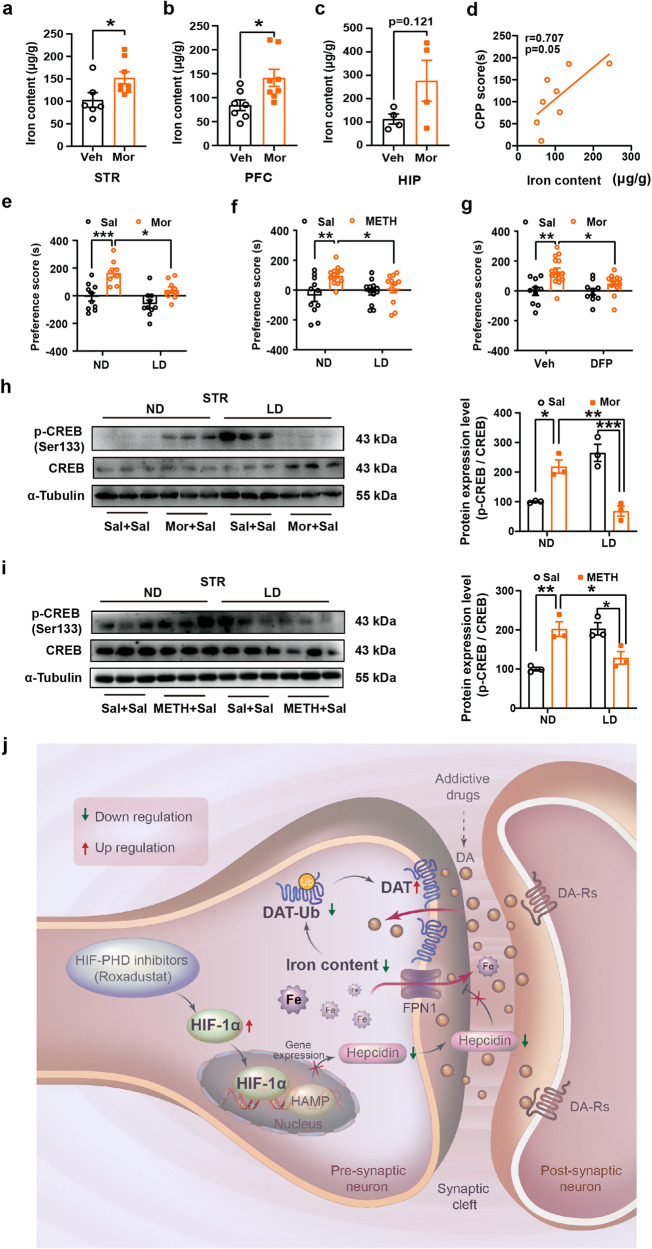
Substance use disorder remains a major challenge, with an enduring need to identify and evaluate new, translational targets for effective treatment. Here, we report the upregulation of Hypoxia-inducible factor-1α (HIF-1α) expression by roxadustat (Rox), a drug developed for renal anemia that inhibits HIF prolyl hydroxylase to prevent degradation of HIF-1α, administered either systemically or locally into selected brain regions, suppressed morphine (Mor)-induced conditioned place preference (CPP). A similar effect was observed with methamphetamine (METH). Moreover, Rox also inhibited the expression of both established and reinstated Mor-CPP and promoted the extinction of Mor-CPP. Additionally, the elevation of HIF-1α enhanced hepcidin/ferroportin 1 (FPN1)-mediated iron efflux and resulted in cellular iron deficiency, which led to the functional accumulation of the dopamine transporter (DAT) in plasma membranes due to iron deficiency-impaired ubiquitin degradation. Notably, iron-deficient mice generated via a low iron diet mimicked the effect of Rox on the prevention of Mor- or METH-CPP formation, without affecting other types of memory. These data reveal a novel mechanism for HIF-1α and iron involvement in substance use disorder, which may represent a potential novel therapeutic strategy for the treatment of drug abuse. The findings also repurpose Rox by suggesting a potential new indication for the treatment of substance use disorder.
© 2023. West China Hospital, Sichuan University.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
