

Co-infection of mice with SARS-CoV-2 and Mycobacterium tuberculosis limits early viral replication but does not affect mycobacterial loads
- PMID: 37720210
- PMCID: PMC10502726
- DOI: 10.3389/fimmu.2023.1240419
Co-infection of mice with SARS-CoV-2 and Mycobacterium tuberculosis limits early viral replication but does not affect mycobacterial loads
Abstract
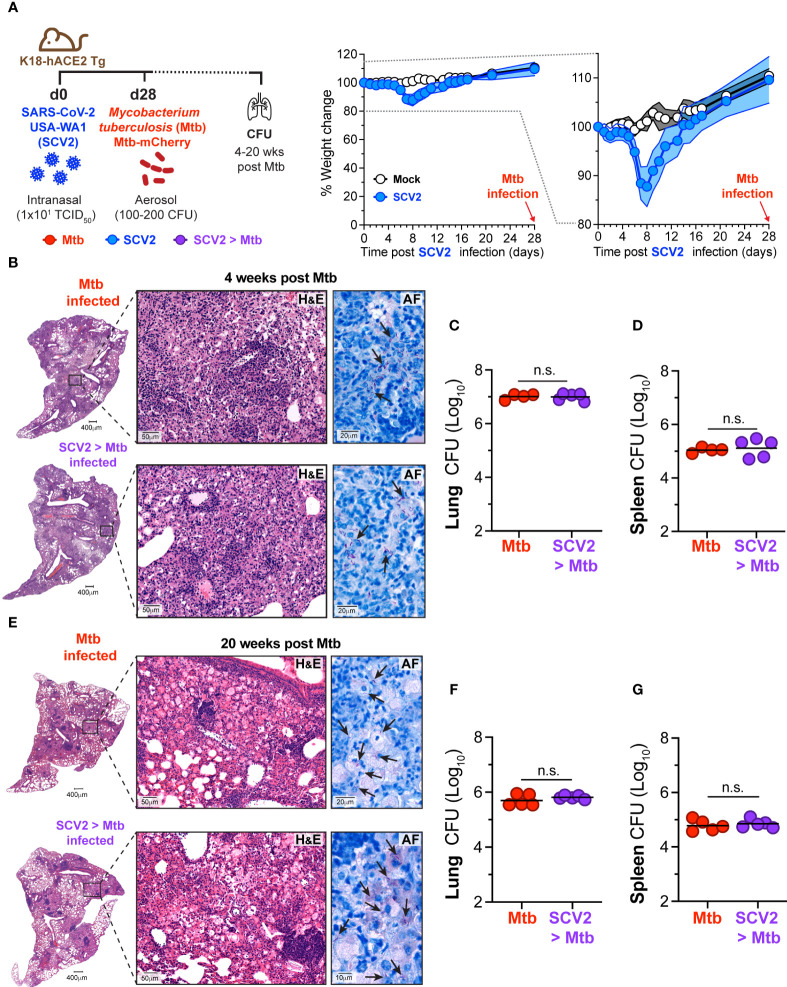
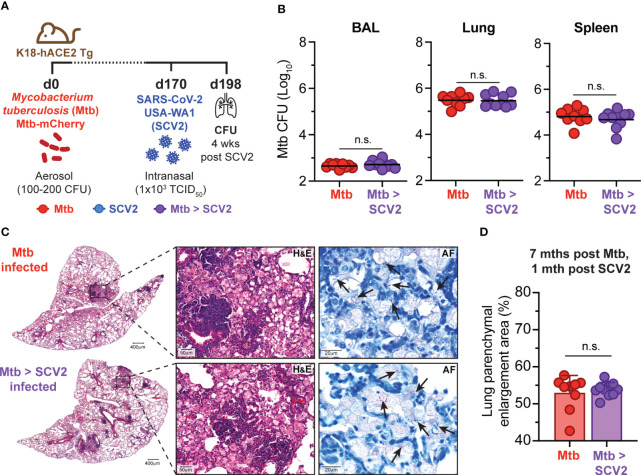
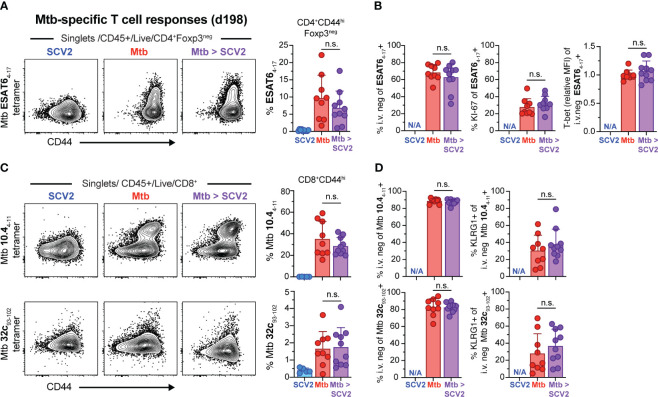
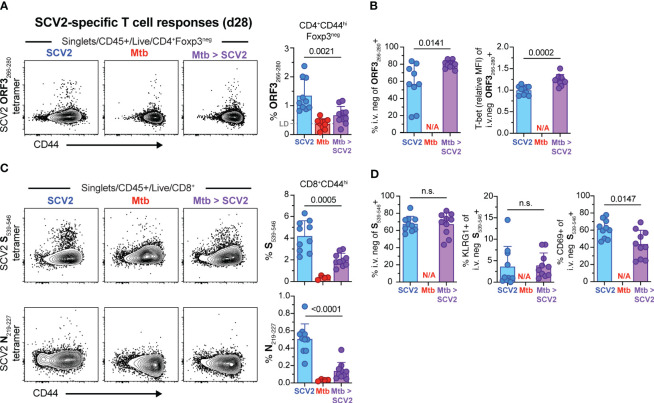
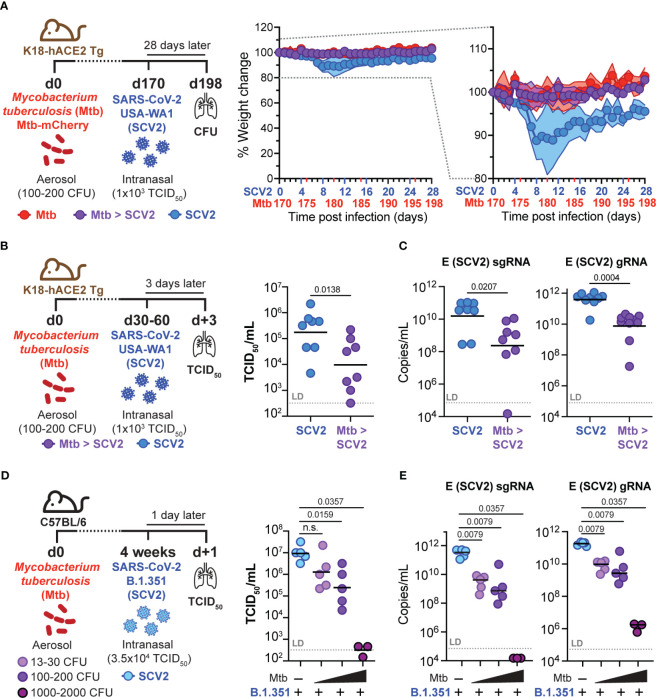
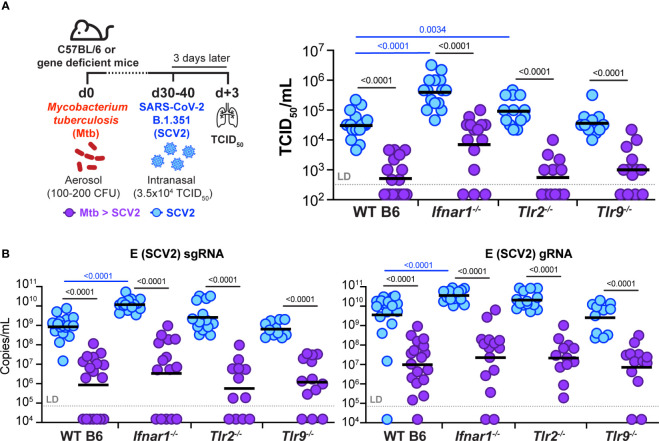
Viral co-infections have been implicated in worsening tuberculosis (TB) and during the COVID-19 pandemic, the global rate of TB-related deaths has increased for the first time in over a decade. We and others have previously shown that a resolved prior or concurrent influenza A virus infection in Mycobacterium tuberculosis (Mtb)-infected mice resulted in increased pulmonary bacterial burden, partly through type I interferon (IFN-I)-dependent mechanisms. Here we investigated whether SARS-CoV-2 (SCV2) co-infection could also negatively affect bacterial control of Mtb. Importantly, we found that K18-hACE2 transgenic mice infected with SCV2 one month before, or months after aerosol Mtb exposure did not display exacerbated Mtb infection-associated pathology, weight loss, nor did they have increased pulmonary bacterial loads. However, pre-existing Mtb infection at the time of exposure to the ancestral SCV2 strain in infected K18-hACE2 transgenic mice or the beta variant (B.1.351) in WT C57Bl/6 mice significantly limited early SCV2 replication in the lung. Mtb-driven protection against SCV2 increased with higher bacterial doses and did not require IFN-I, TLR2 or TLR9 signaling. These data suggest that SCV2 co-infection does not exacerbate Mtb infection in mice, but rather the inflammatory response generated by Mtb infection in the lungs at the time of SCV2 exposure restricts viral replication.
Keywords: COVID-19; SARS-CoV-2; Type-I interferon; co-infection; lung; mycobacterium tuberculosis; tuberculosis.
Copyright © 2023 Baker, Amaral, Castro, Bohrer, Torres-Juárez, Jordan, Nelson, Barber, Johnson, Hilligan and Mayer-Barber.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous