

Multiplexed Assessment of Promiscuous Non-Canonical Amino Acid Synthase Activity in a Pyridoxal Phosphate-Dependent Protein Family
- PMID: 37720819
- PMCID: PMC10501158
- DOI: 10.1021/acscatal.3c02498
Multiplexed Assessment of Promiscuous Non-Canonical Amino Acid Synthase Activity in a Pyridoxal Phosphate-Dependent Protein Family
Abstract
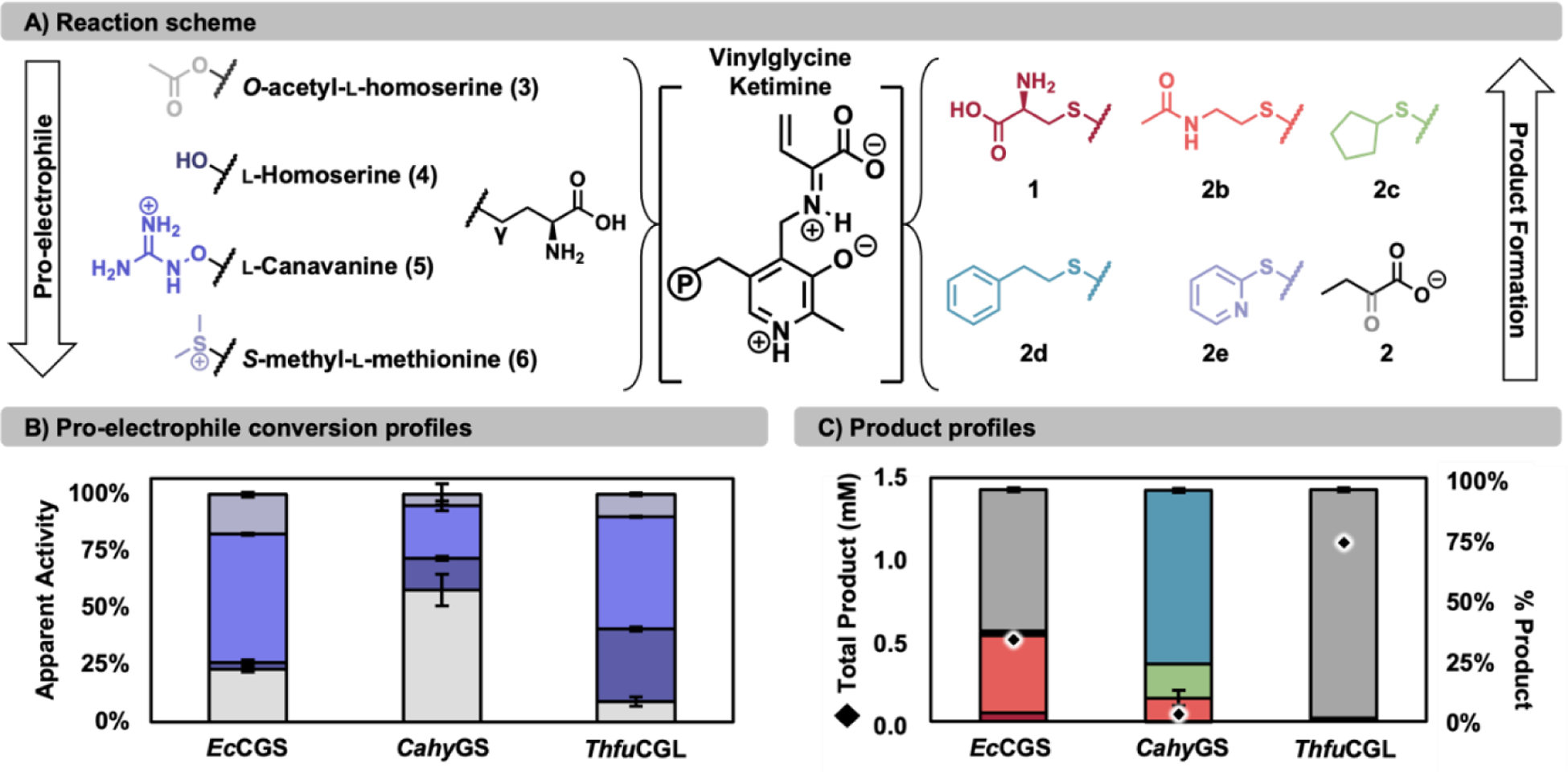
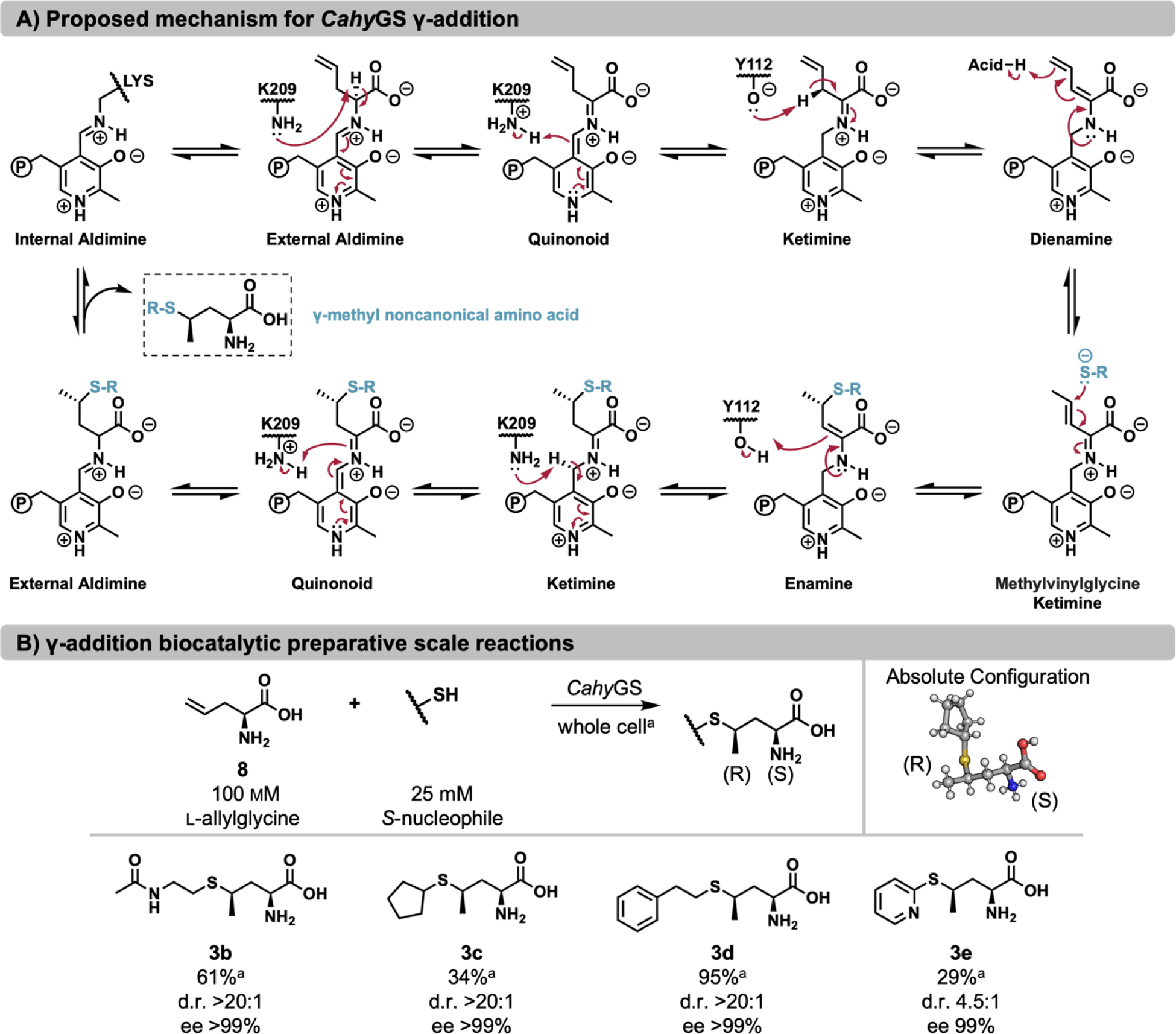
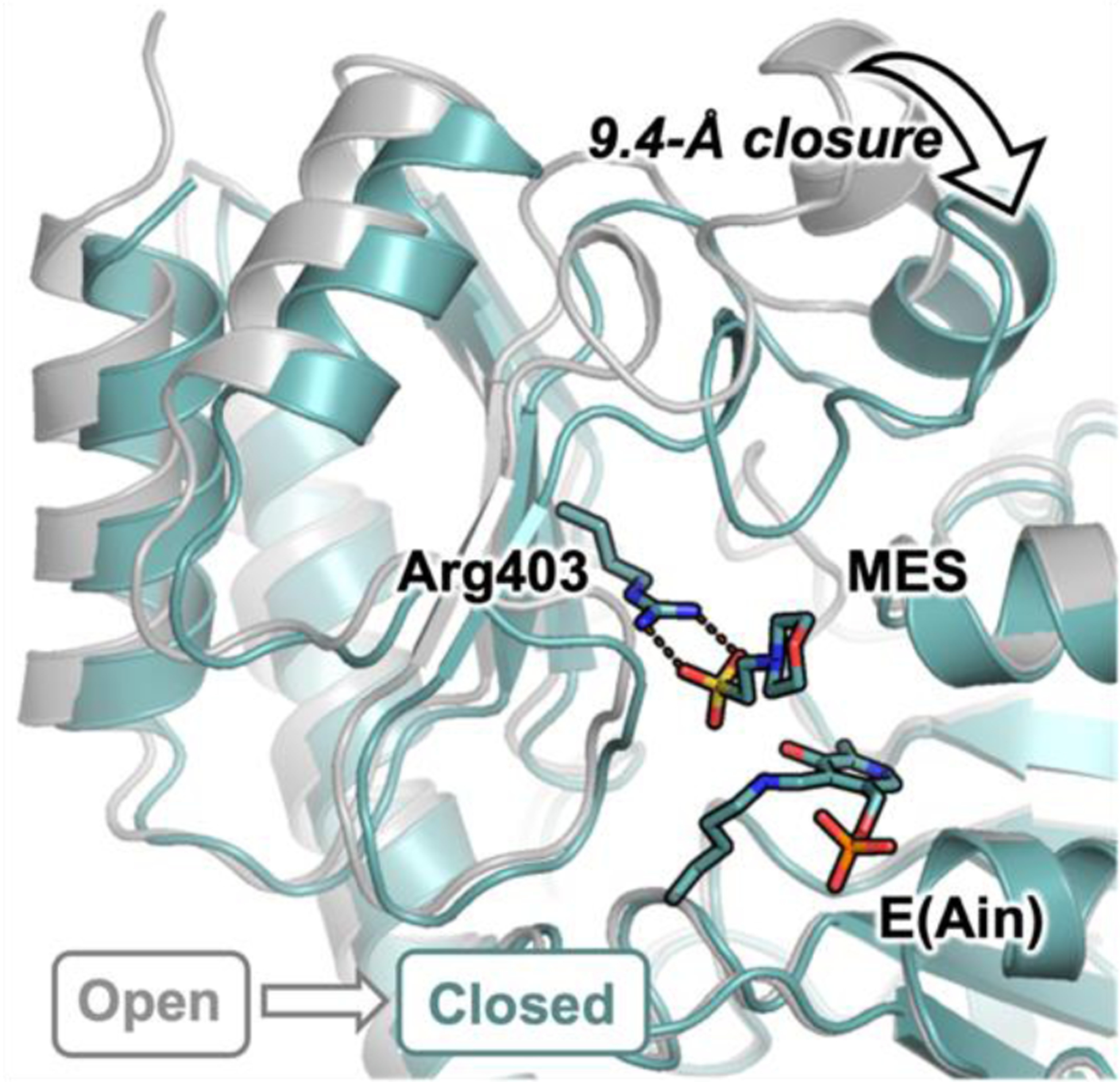
Pyridoxal phosphate (PLP)-dependent enzymes afford access to a variety of non-canonical amino acids (ncAAs), which are premier buildings blocks for the construction of complex bioactive molecules. The vinylglycine ketimine (VGK) subfamily of PLP-dependent enzymes plays a critical role in sulfur metabolism and is home to a growing set of secondary metabolic enzymes that synthesize γ-substituted ncAAs. Identification of VGK enzymes for biocatalysis faces a distinct challenge because the subfamily contains both desirable synthases as well as lyases that break down ncAAs. Some enzymes have both activities, which may contribute to pervasive mis-annotation. To navigate this complex functional landscape, we used a substrate multiplexed screening approach to rapidly measure the substrate promiscuity of 40 homologs in the VGK subfamily. We found that enzymes involved in transsulfuration are less likely to have promiscuous activities and often possess undesirable lyase activity. Enzymes from direct sulfuration and secondary metabolism generally had a high degree of substrate promiscuity. From this cohort, we identified an exemplary γ-synthase from Caldicellulosiruptor hydrothermalis (CahyGS). This enzyme is thermostable and has high expression (~400 mg protein per L culture), enabling preparative scale synthesis of thioether containing ncAAs. When assayed with l-allylglycine, CahyGS catalyzes a stereoselective γ-addition reaction to afford access to a unique set of γ-methyl branched ncAAs. We determined high-resolution crystal structures of this enzyme that define an open-close transition associated with ligand binding and set the stage for future engineering within this enzyme subfamily.
Keywords: Biocatalysis; competition reactions; methionine analogs; noncanonical amino acids; sequence similarity network analysis.
Figures





Similar articles
-
The enzymes of the transsulfuration pathways: active-site characterizations.Biochim Biophys Acta. 2011 Nov;1814(11):1511-7. doi: 10.1016/j.bbapap.2011.03.006. Epub 2011 Mar 22. Biochim Biophys Acta. 2011. PMID: 21435402 Review.
-
Discovery and Biocatalytic Application of a PLP-Dependent Amino Acid γ-Substitution Enzyme That Catalyzes C-C Bond Formation.J Am Chem Soc. 2020 Jun 10;142(23):10506-10515. doi: 10.1021/jacs.0c03535. Epub 2020 Jun 1. J Am Chem Soc. 2020. PMID: 32434326 Free PMC article.
-
Engineering enzymes for noncanonical amino acid synthesis.Chem Soc Rev. 2018 Dec 21;47(24):8980-8997. doi: 10.1039/c8cs00665b. Epub 2018 Oct 3. Chem Soc Rev. 2018. PMID: 30280154 Free PMC article. Review.
-
Engineered Biocatalytic Synthesis of β-N-Substituted-α-Amino Acids.Angew Chem Int Ed Engl. 2023 Oct 23;62(43):e202311189. doi: 10.1002/anie.202311189. Epub 2023 Sep 14. Angew Chem Int Ed Engl. 2023. PMID: 37625129 Free PMC article.
-
Diverse functional evolution of serine decarboxylases: identification of two novel acetaldehyde synthases that uses hydrophobic amino acids as substrates.BMC Plant Biol. 2014 Sep 18;14:247. doi: 10.1186/s12870-014-0247-x. BMC Plant Biol. 2014. PMID: 25230835 Free PMC article.
Cited by
-
Discovery and Characterization of Pyridoxal 5'-Phosphate-Dependent Cycloleucine Synthases.J Am Chem Soc. 2024 May 29;146(21):14672-14684. doi: 10.1021/jacs.4c02142. Epub 2024 May 14. J Am Chem Soc. 2024. PMID: 38743881 Free PMC article.
-
Elucidation of the stereochemical mechanism of cystathionine γ-lyase reveals how substrate specificity constrains catalysis.ACS Catal. 2024 Aug 2;14(15):11196-11204. doi: 10.1021/acscatal.4c02281. Epub 2024 Jul 11. ACS Catal. 2024. PMID: 39391268 Free PMC article.
-
Chemoenzymatic Cascade Synthesis of Metal-Chelating α-Amino Acids.ChemCatChem. 2025 Mar 6;17(5):e202401958. doi: 10.1002/cctc.202401958. Epub 2024 Dec 9. ChemCatChem. 2025. PMID: 40761323 Free PMC article.
-
An efficient lysate-based approach for biosynthesis of the pyrrolobenzodiazepine natural product tilimycin.J Biotechnol. 2025 Jun;402:87-95. doi: 10.1016/j.jbiotec.2025.03.012. Epub 2025 Mar 20. J Biotechnol. 2025. PMID: 40120764
-
Molecular Basis for Cγ-N Bond Formation by PLP-Dependent Enzyme LolC.Biochemistry. 2024 Dec 17;63(24):3348-3356. doi: 10.1021/acs.biochem.4c00588. Epub 2024 Dec 6. Biochemistry. 2024. PMID: 39641520 Free PMC article.
References
-
- Savile CK; Janey JM; Mundorff EC; Moore JC; Tam S; Jarvis WR; Colbeck JC; Krebber A; Fleitz FJ; Brands J; Devine PN; Huisman GW; Hughes GJ Biocatalytic Asymmetric Synthesis of Chiral Amines from Ketones Applied to Sitagliptin Manufacture. Science 2010, 329 (5989), 305–309. 10.1126/science.1188934. - DOI - PubMed
-
- Rocha JF; Pina AF; Sousa SF; Cerqueira NMFSA PLP-Dependent Enzymes as Important Biocatalysts for the Pharmaceutical{,} Chemical and Food Industries: A Structural and Mechanistic Perspective. Catal. Sci. Technol 2019, 9 (18), 4864–4876. 10.1039/C9CY01210A. - DOI
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials