

Friction patterns guide actin network contraction
- PMID: 37725653
- PMCID: PMC10523593
- DOI: 10.1073/pnas.2300416120
Friction patterns guide actin network contraction
Abstract
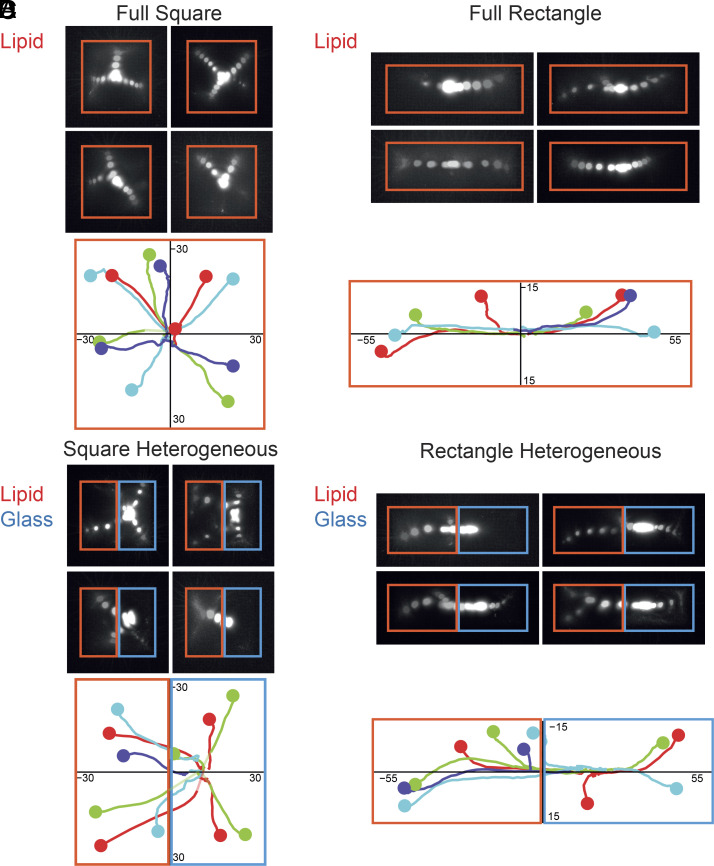
The shape of cells is the outcome of the balance of inner forces produced by the actomyosin network and the resistive forces produced by cell adhesion to their environment. The specific contributions of contractile, anchoring and friction forces to network deformation rate and orientation are difficult to disentangle in living cells where they influence each other. Here, we reconstituted contractile actomyosin networks in vitro to study specifically the role of the friction forces between the network and its anchoring substrate. To modulate the magnitude and spatial distribution of friction forces, we used glass or lipids surface micropatterning to control the initial shape of the network. We adapted the concentration of Nucleating Promoting Factor on each surface to induce the assembly of actin networks of similar densities and compare the deformation of the network toward the centroid of the pattern shape upon myosin-induced contraction. We found that actin network deformation was faster and more coordinated on lipid bilayers than on glass, showing the resistance of friction to network contraction. To further study the role of the spatial distribution of these friction forces, we designed heterogeneous micropatterns made of glass and lipids. The deformation upon contraction was no longer symmetric but biased toward the region of higher friction. Furthermore, we showed that the pattern of friction could robustly drive network contraction and dominate the contribution of asymmetric distributions of myosins. Therefore, we demonstrate that during contraction, both the active and resistive forces are essential to direct the actin network deformation.
Keywords: actin; contraction; cytoskeleton; friction.
Conflict of interest statement
The authors declare no competing interest.
Figures





References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
