

CD1 lipidomes reveal lipid-binding motifs and size-based antigen-display mechanisms
- PMID: 37725977
- PMCID: PMC10591967
- DOI: 10.1016/j.cell.2023.08.022
CD1 lipidomes reveal lipid-binding motifs and size-based antigen-display mechanisms
Abstract
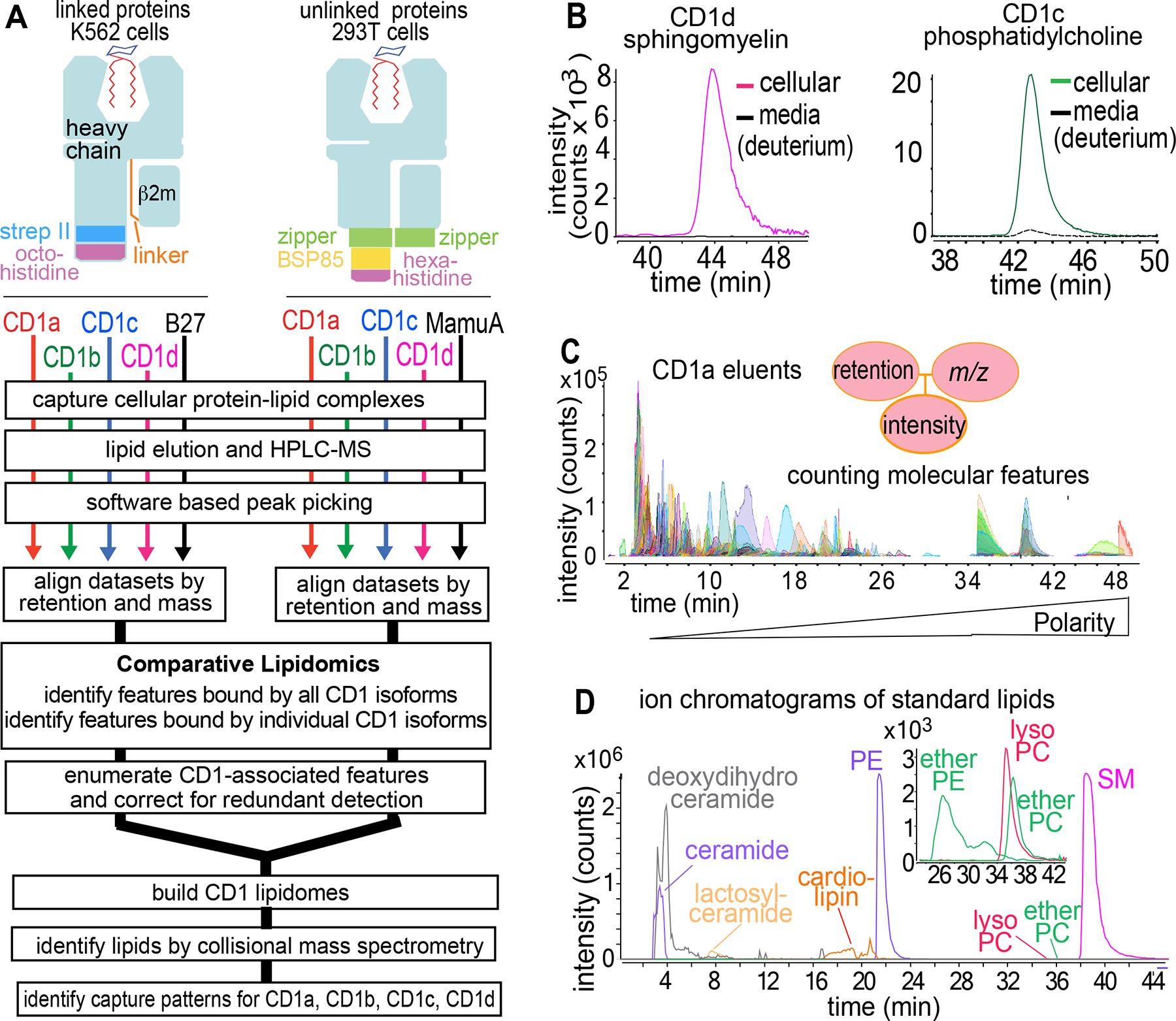
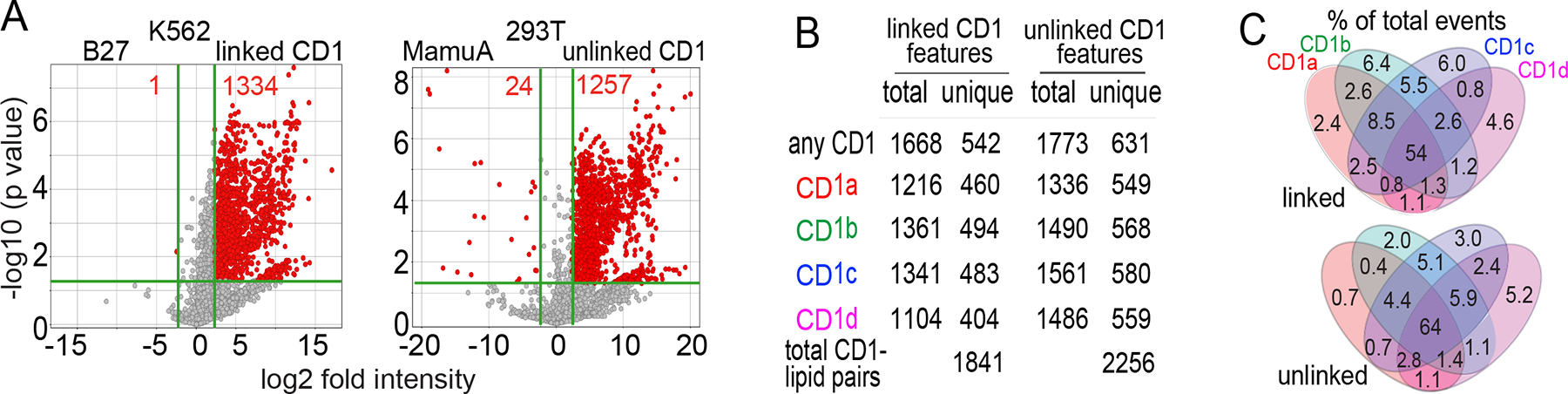
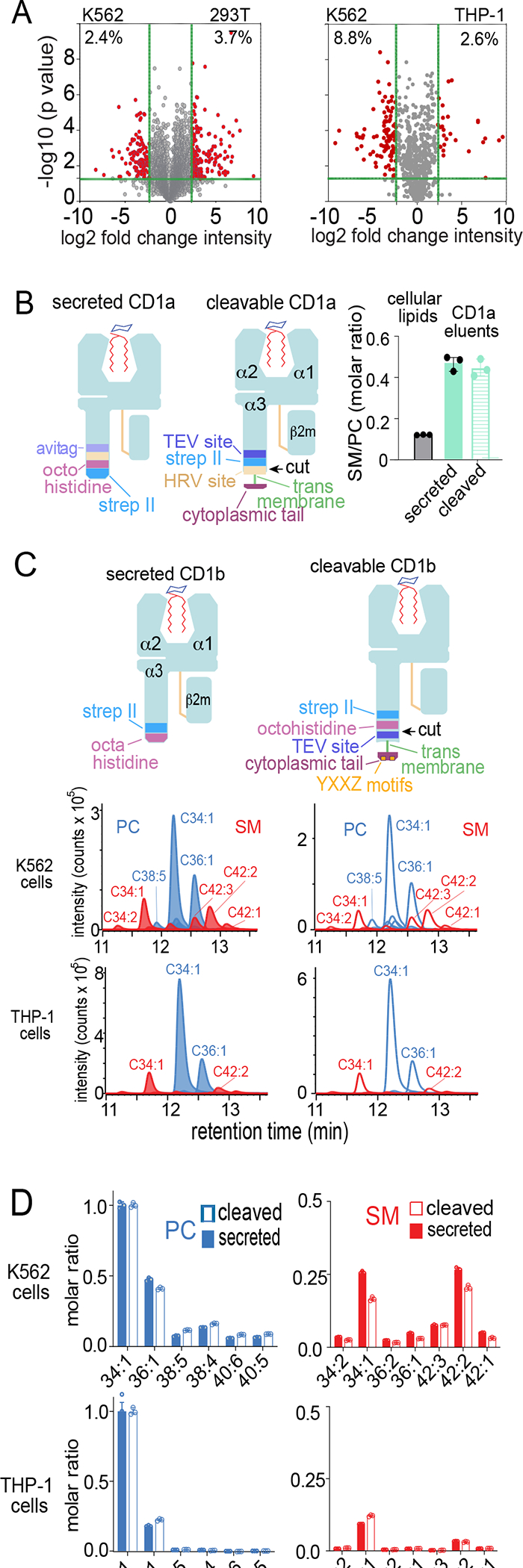
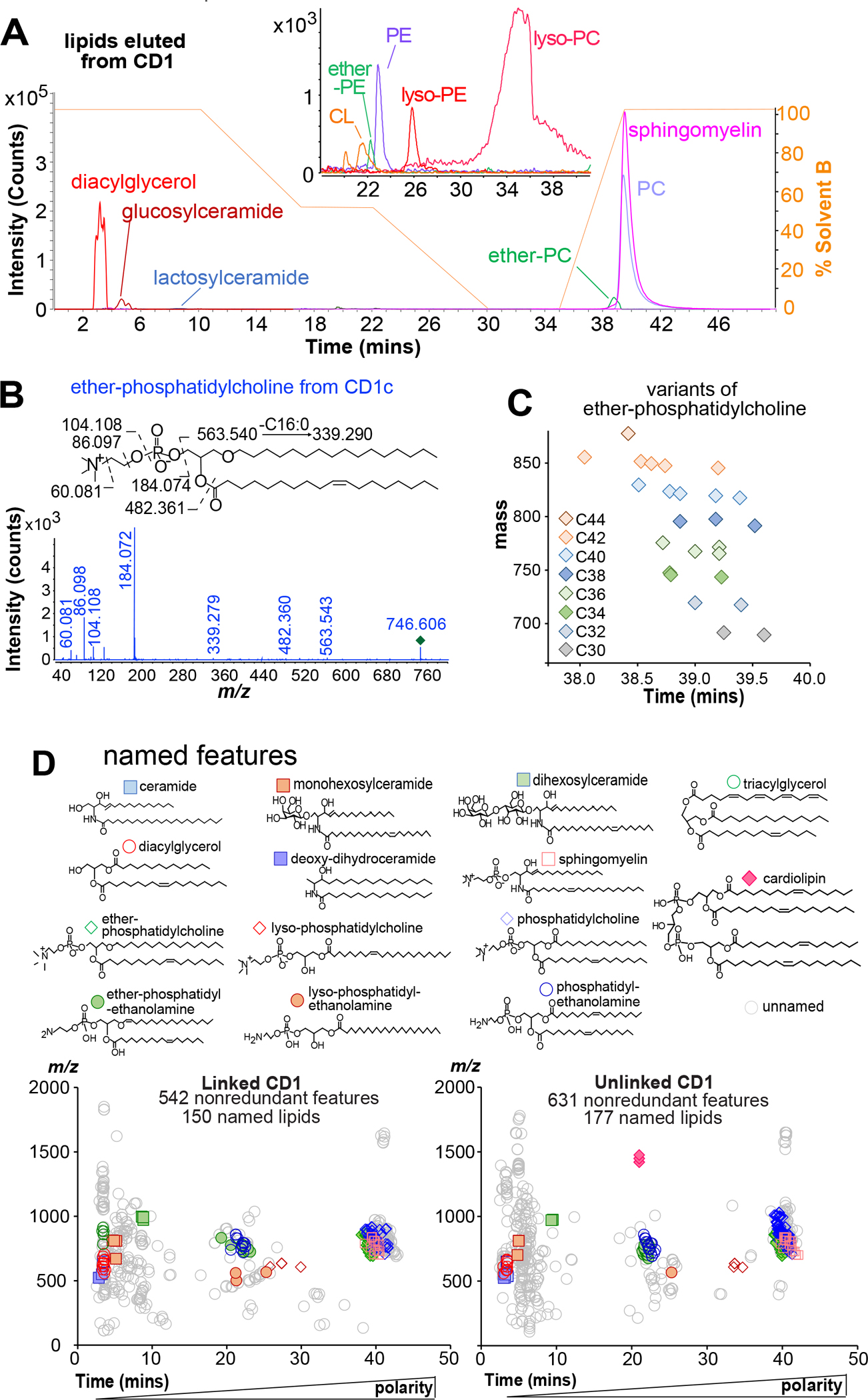
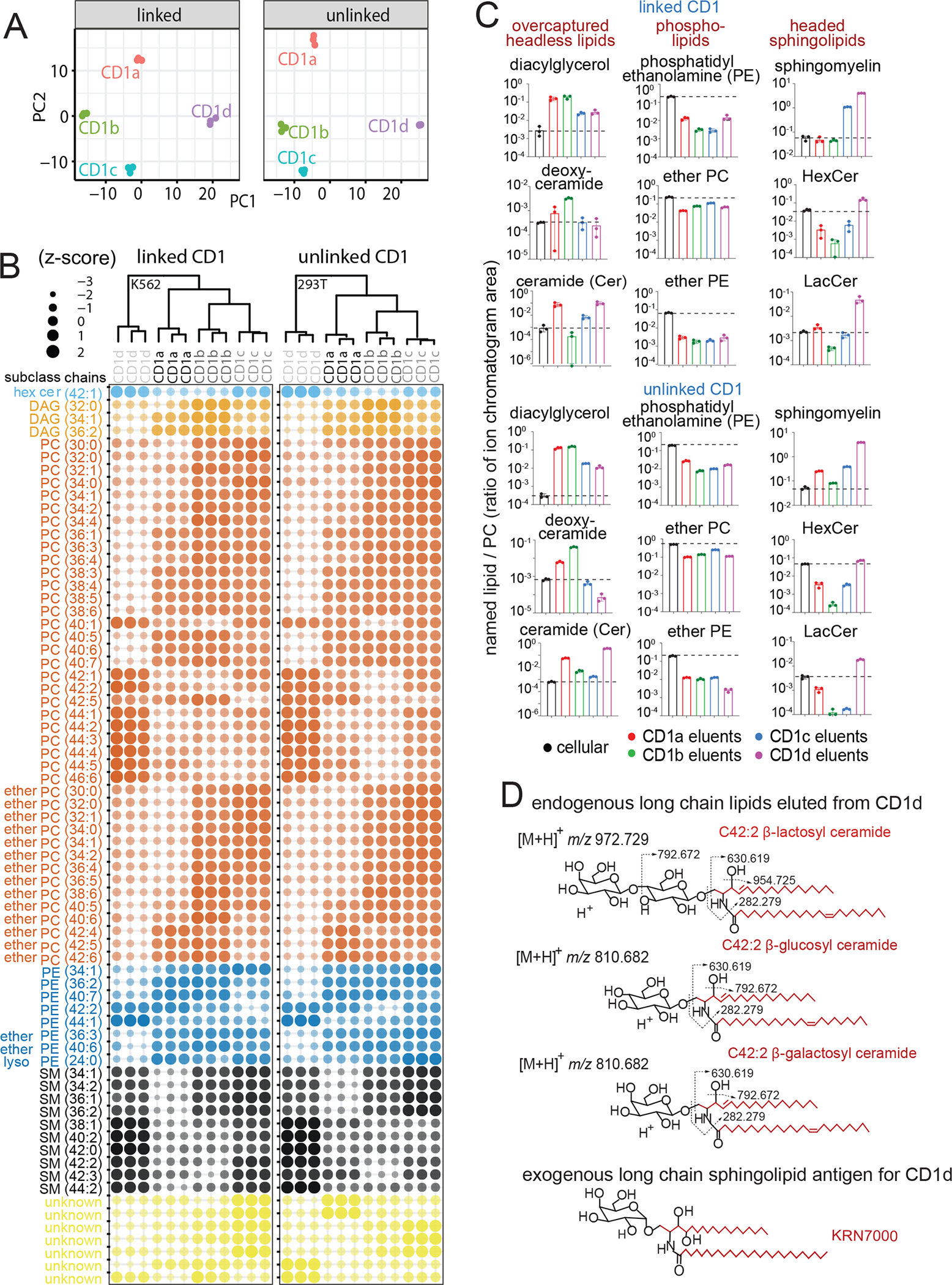
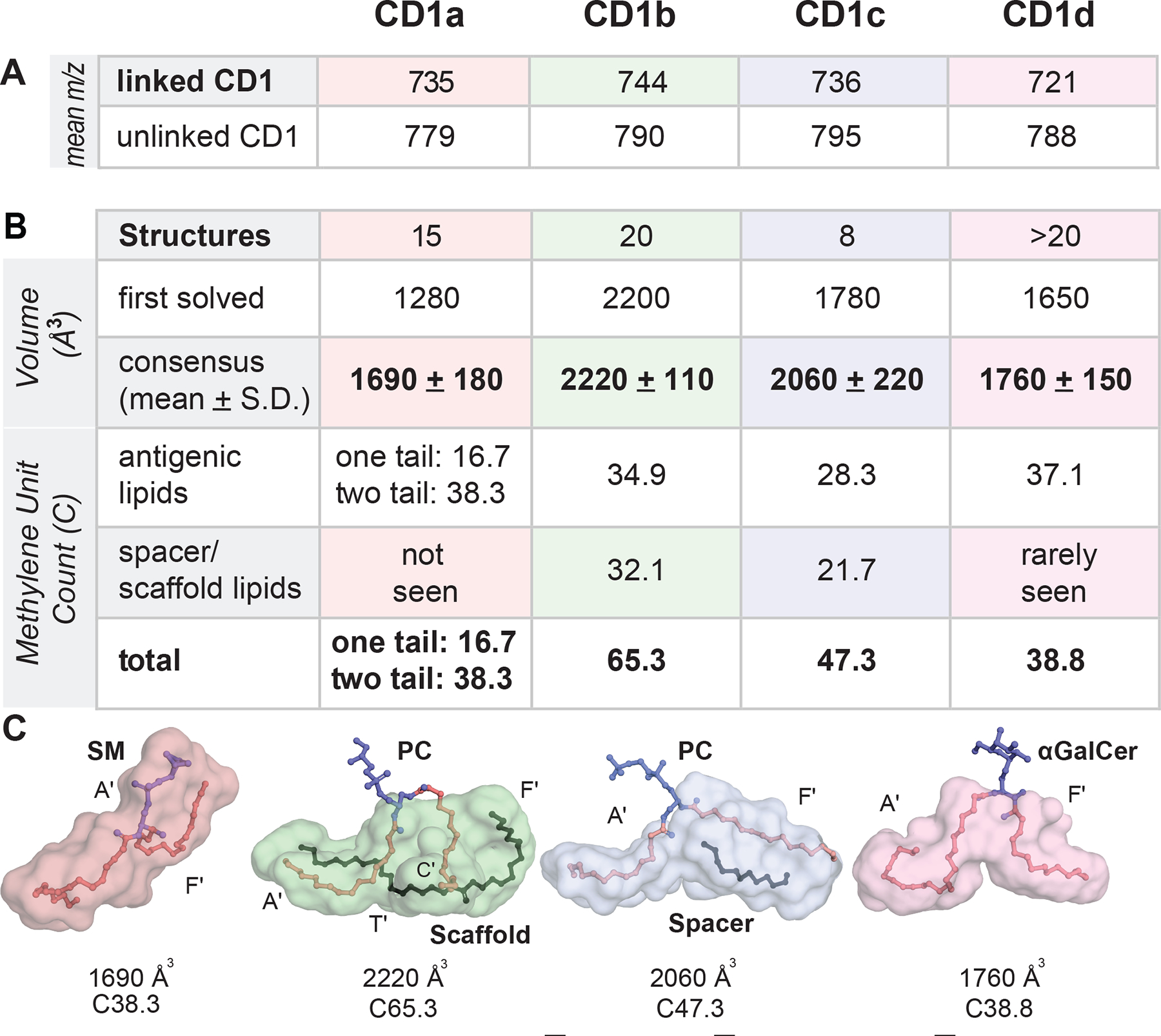
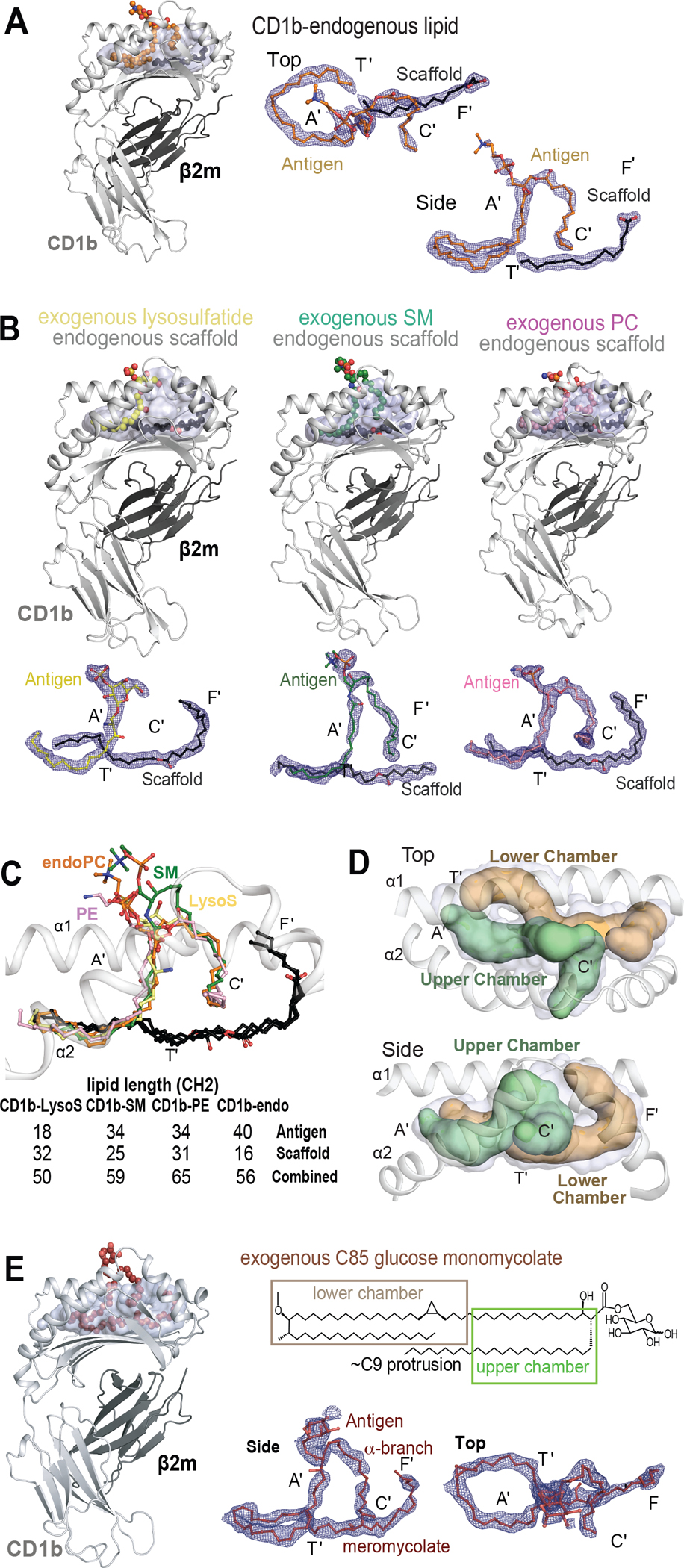
The CD1 system binds lipid antigens for display to T cells. Here, we solved lipidomes for the four human CD1 antigen-presenting molecules, providing a map of self-lipid display. Answering a basic question, the detection of >2,000 CD1-lipid complexes demonstrates broad presentation of self-sphingolipids and phospholipids. Whereas peptide antigens are chemically processed, many lipids are presented in an unaltered form. However, each type of CD1 protein differentially edits the self-lipidome to show distinct capture motifs based on lipid length and chemical composition, suggesting general antigen display mechanisms. For CD1a and CD1d, lipid size matches the CD1 cleft volume. CD1c cleft size is more variable, and CD1b is the outlier, where ligands and clefts show an extreme size mismatch that is explained by uniformly seating two small lipids in one cleft. Furthermore, the list of compounds that comprise the integrated CD1 lipidome supports the ongoing discovery of lipid blockers and antigens for T cells.
Keywords: CD1; T cell receptor; T cells; antigen presentation; antigen processing; dendritic cells; lipidomics; major histocompatibility complex; sphingolipids.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors hold intellectual property through the Massachusetts General Brigham (D.B.M., G.S.O., and J.R.; filing 29618-0390P01) and the University of Oxford (G.S.O., C.S.H., and Y.-L.C.; filings 2116709.3, 2217923.8, 2217924.6). G.S.O. collaborates with UCB and Janssen, and D.B.M. consults for Pfizer.
Figures
References
-
- Garboczi DN, Ghosh P, Utz U, Fan QR, Biddison WE, Wiley, and DC (1996). Structure of the complex between human T-cell receptor, viral peptide and HLA-A2. Nature 384, 134–141. - PubMed
-
- Garcia KC, Degano M, Stanfield RL, Brunmark A, Jackson MR, Peterson PA, Teyton L, and Wilson IA (1996). An alphabeta T cell receptor structure at 2.5 A and its orientation in the TCR-MHC complex. Science 274, 209–219. - PubMed
-
- Beckman EM, Porcelli SA, Morita CT, Behar SM, Furlong ST, and Brenner MB (1994). Recognition of a lipid antigen by CD1-restricted àá T cells. Nature 372, 691–694. - PubMed
-
- Zeng Z, Casta AR, Segelke BW, Stura EA, Peterson PA, and Wilson IA. (1997). Crystal structure of mouse CD1: an MHC-like fold with a large hydrophobic binding groove. Science 277, 339–345. - PubMed
-
- Fowlkes BJ, Kruisbeek AM, Ton-That H, Weston MA, Coligan JE, Schwartz RH, and Pardoll DM (1987). A novel population of T-cell receptor alpha beta-bearing thymocytes which predominantly expresses a single V beta gene family. Nature 329, 251–254. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R21 AI115358/AI/NIAID NIH HHS/United States
- 75N92020D00005/HL/NHLBI NIH HHS/United States
- 209222/Z/17/Z/WT_/Wellcome Trust/United Kingdom
- 75N93022D00005/AI/NIAID NIH HHS/United States
- MC_UU_00008/5/MRC_/Medical Research Council/United Kingdom
- 75N95020D00005/DA/NIDA NIH HHS/United States
- 75N99020D00005/OF/ORFDO NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- 75N93023D00005/AI/NIAID NIH HHS/United States
- MC_UU_00008/MRC_/Medical Research Council/United Kingdom
- MC_UU_00008/6/MRC_/Medical Research Council/United Kingdom
- MC_UU_00036/2/MRC_/Medical Research Council/United Kingdom
- R01 AI049313/AI/NIAID NIH HHS/United States
- R01 AR048632/AR/NIAMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
