

Three-dimensional reconstructions of mechanosensory end organs suggest a unifying mechanism underlying dynamic, light touch
- PMID: 37725982
- PMCID: PMC10773061
- DOI: 10.1016/j.neuron.2023.08.023
Three-dimensional reconstructions of mechanosensory end organs suggest a unifying mechanism underlying dynamic, light touch
Abstract
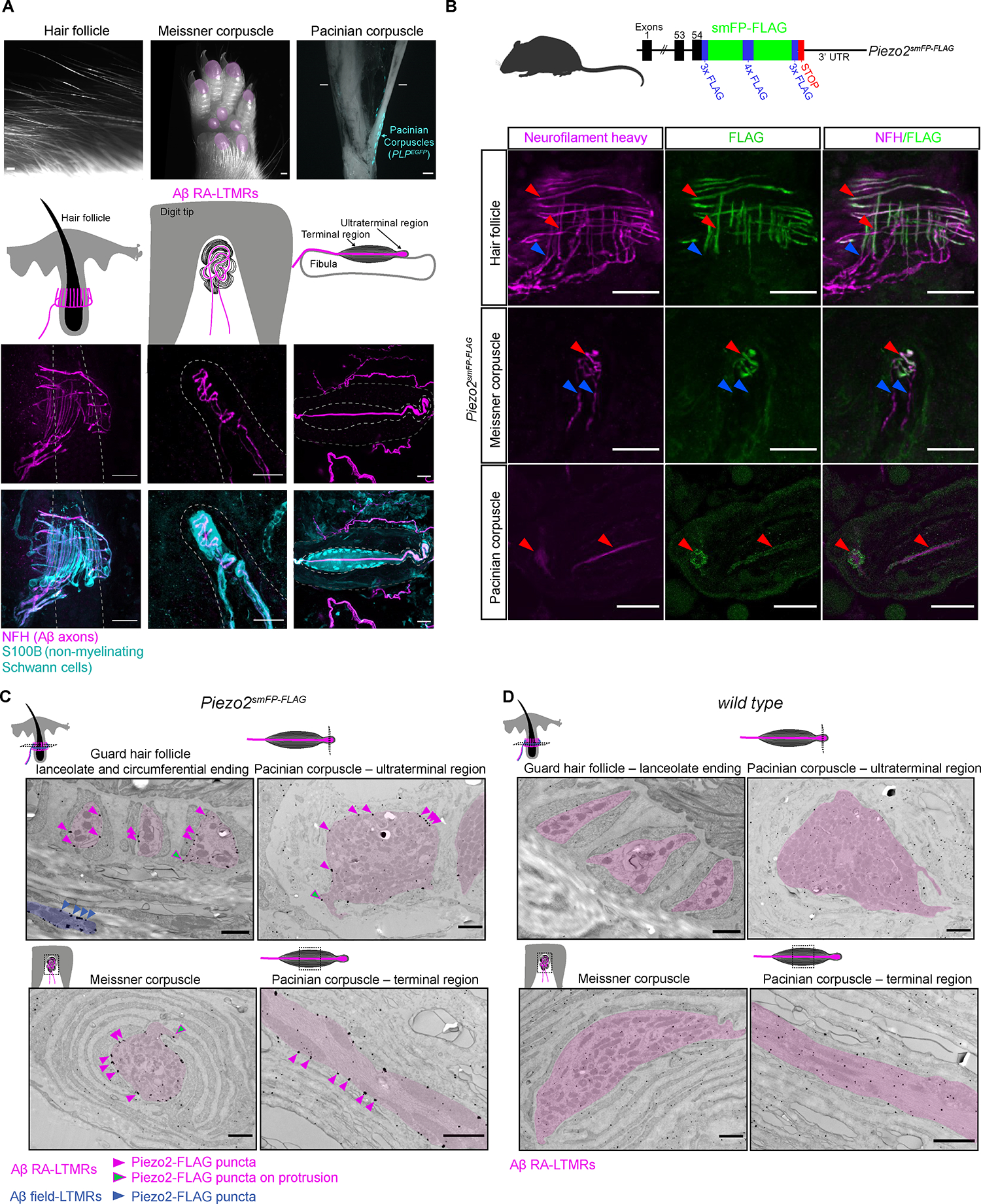
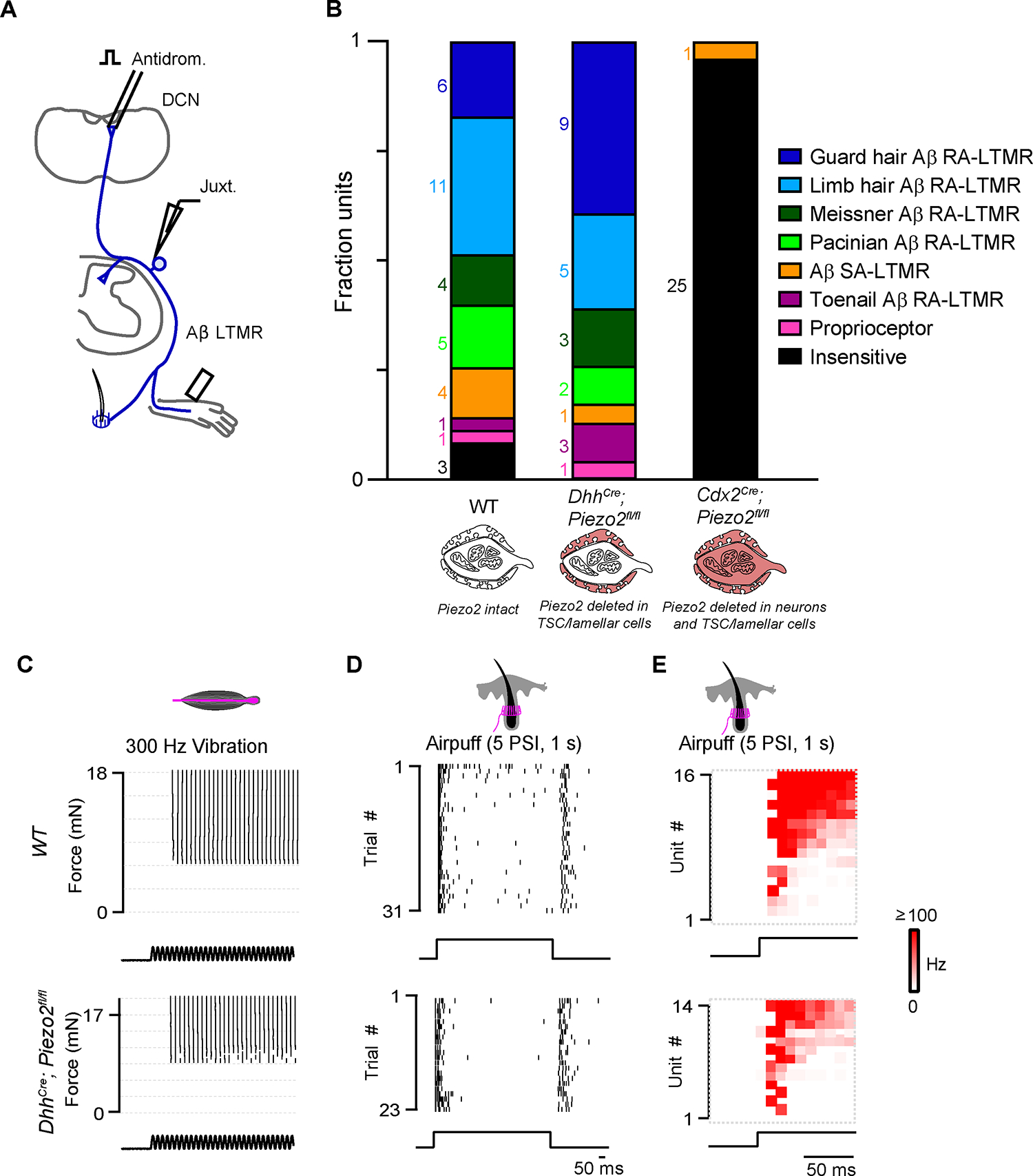
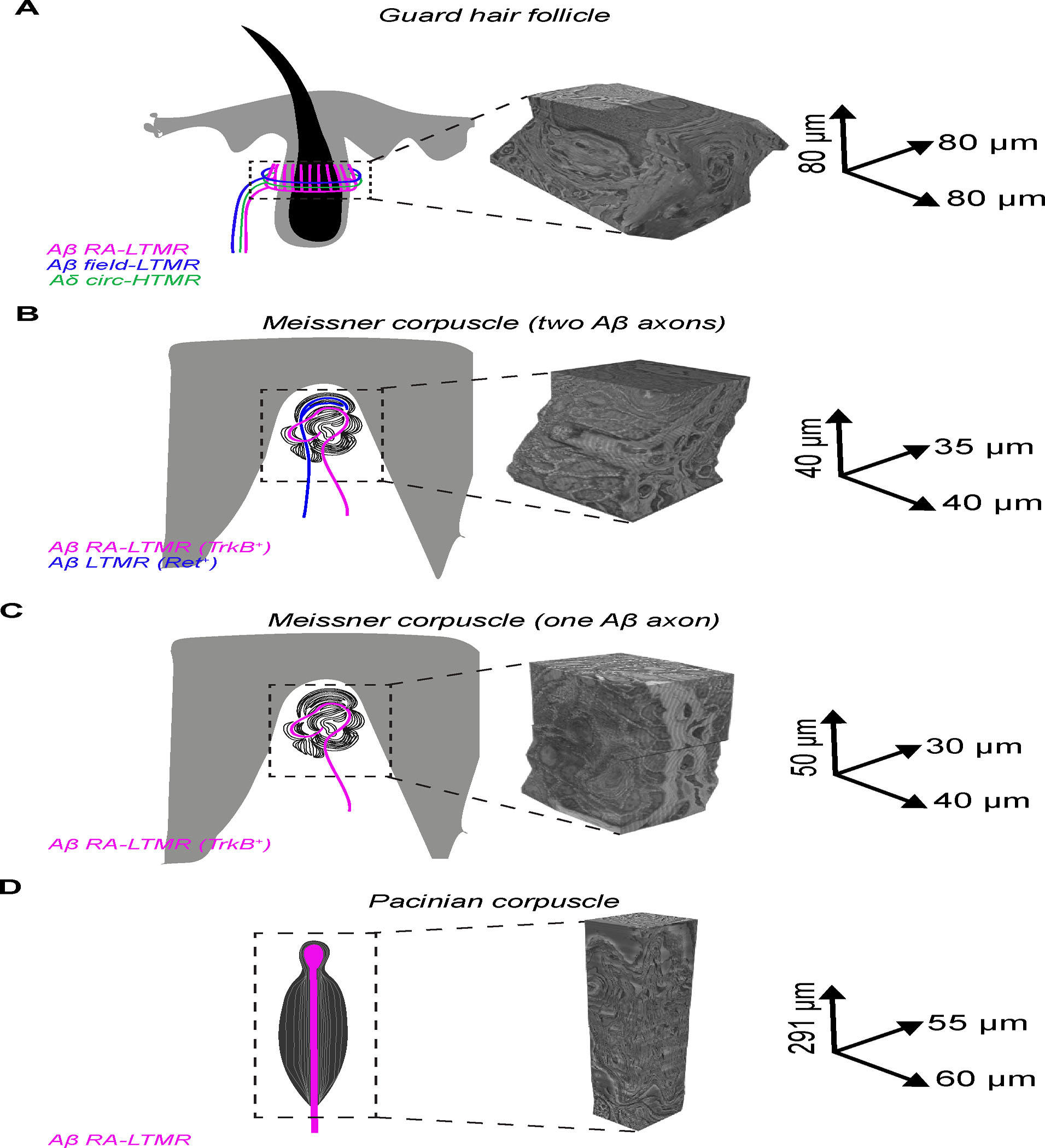
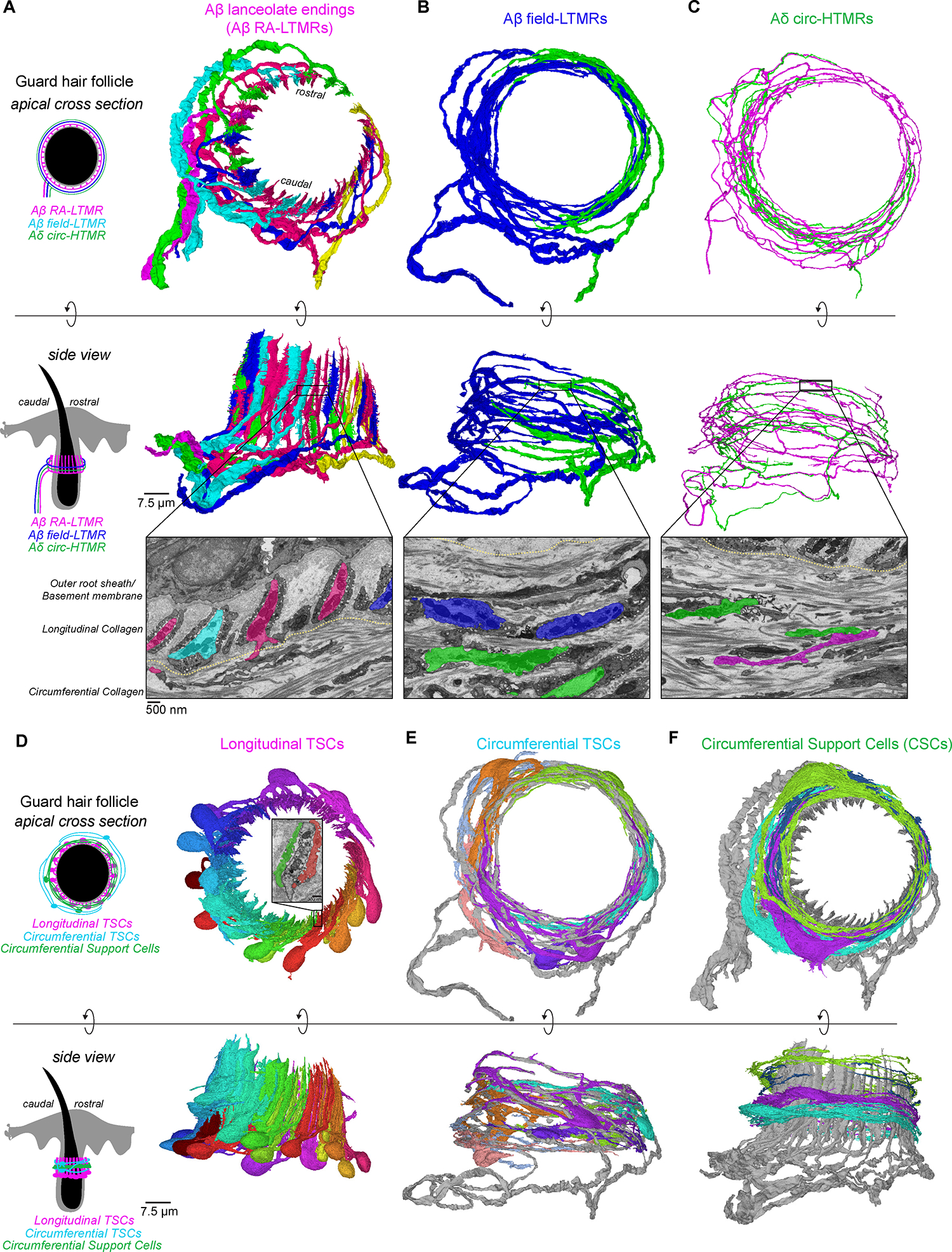
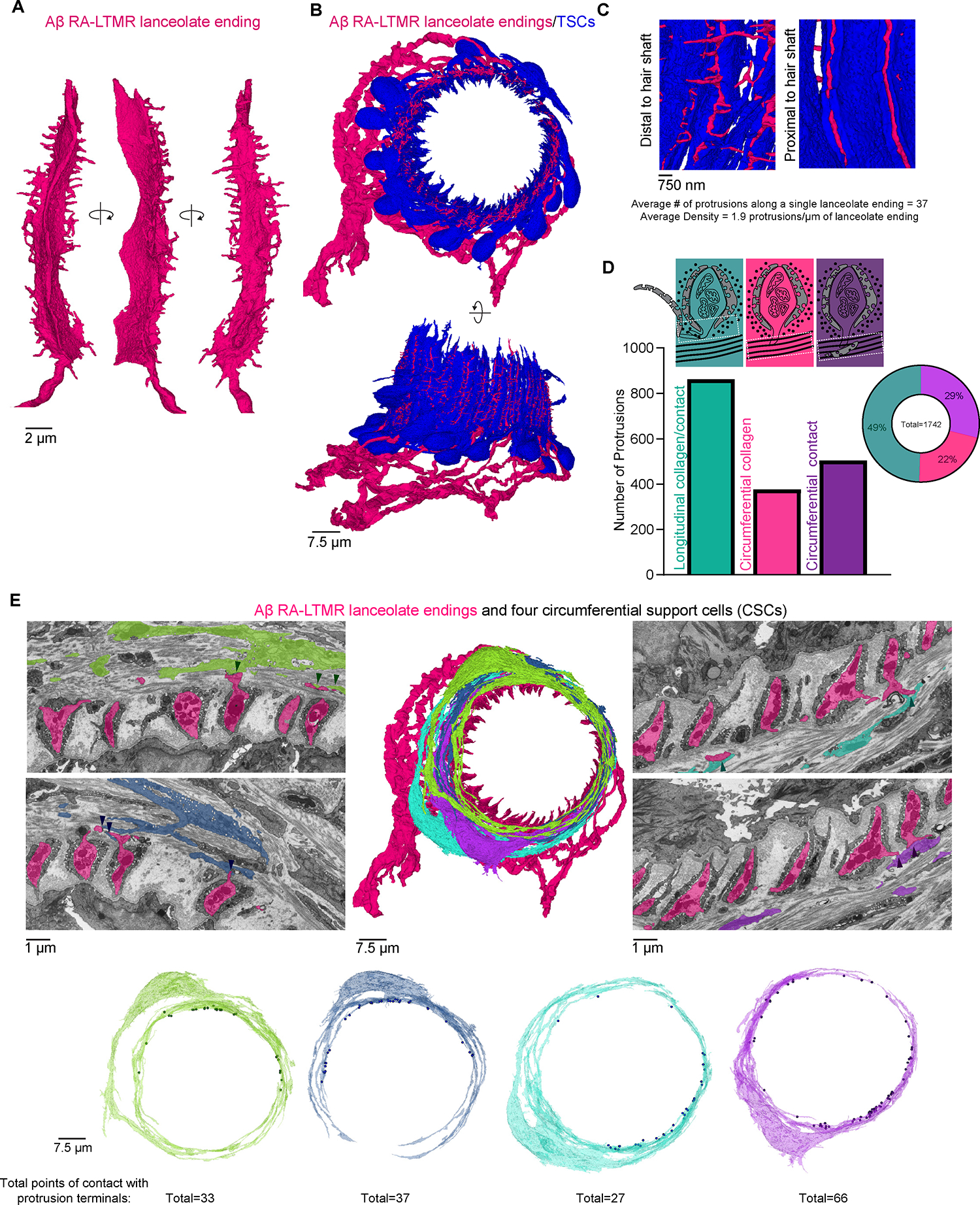
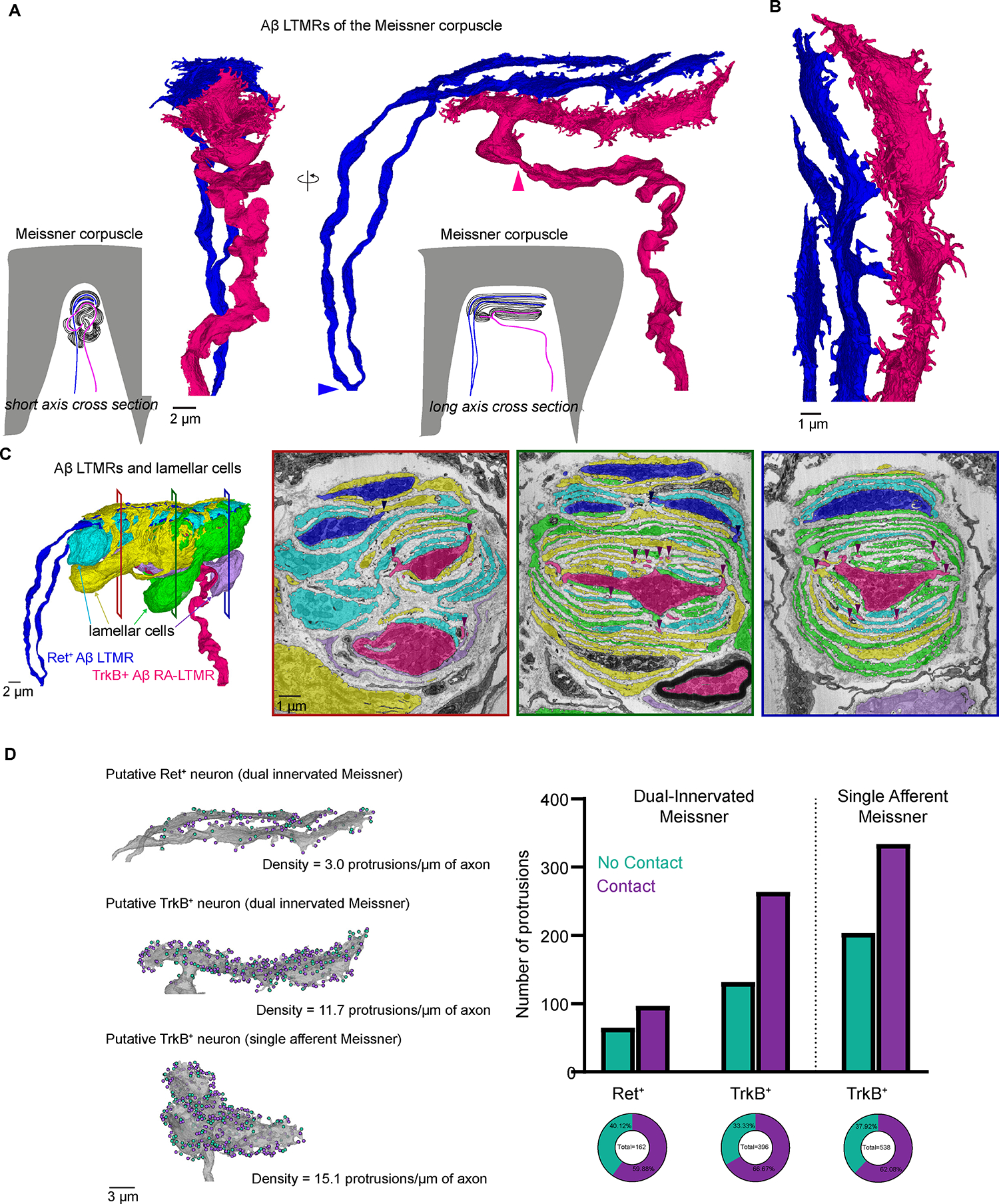
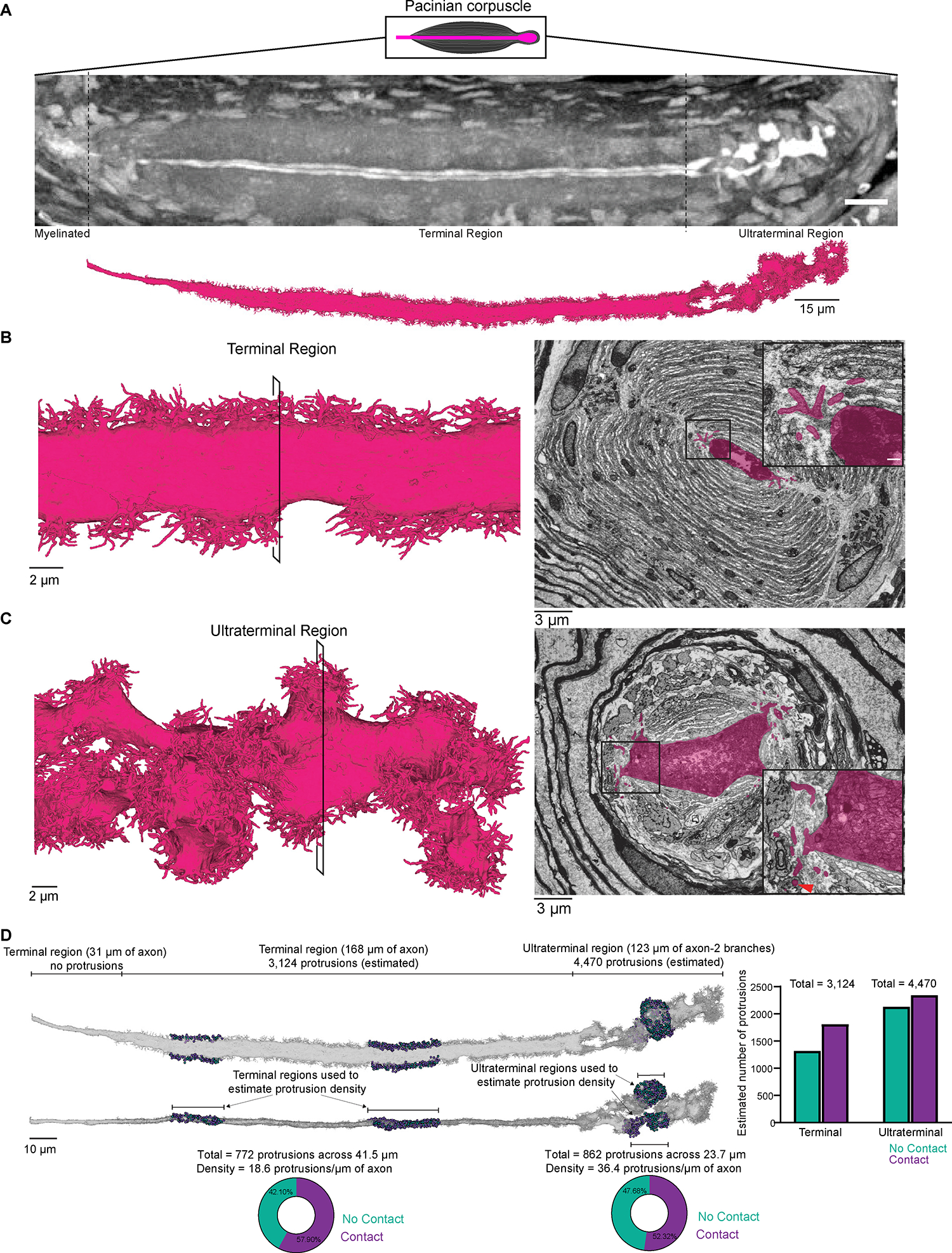
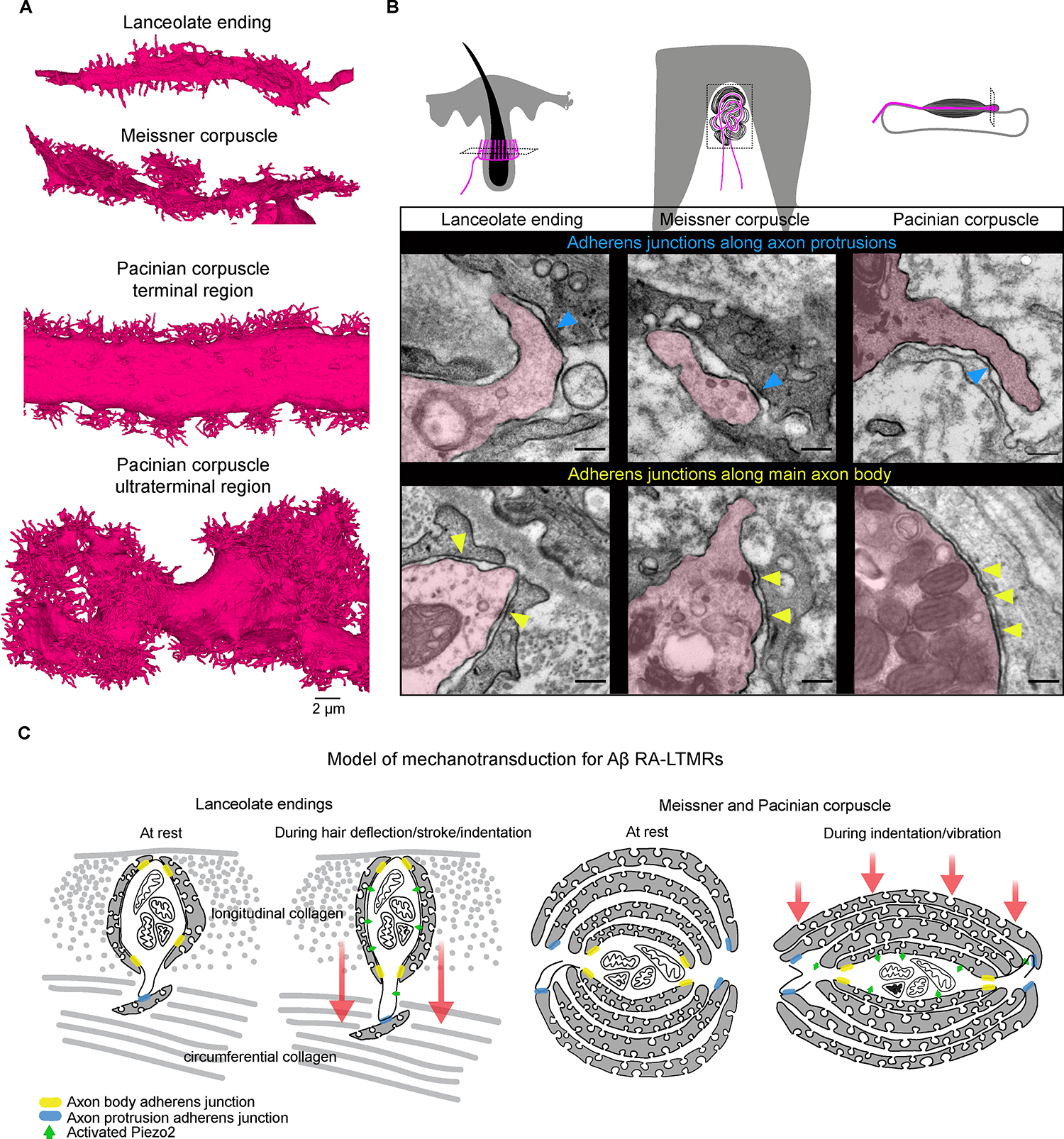
Across mammalian skin, structurally complex and diverse mechanosensory end organs respond to mechanical stimuli and enable our perception of dynamic, light touch. How forces act on morphologically dissimilar mechanosensory end organs of the skin to gate the requisite mechanotransduction channel Piezo2 and excite mechanosensory neurons is not understood. Here, we report high-resolution reconstructions of the hair follicle lanceolate complex, Meissner corpuscle, and Pacinian corpuscle and the subcellular distribution of Piezo2 within them. Across all three end organs, Piezo2 is restricted to the sensory axon membrane, including axon protrusions that extend from the axon body. These protrusions, which are numerous and elaborate extensively within the end organs, tether the axon to resident non-neuronal cells via adherens junctions. These findings support a unified model for dynamic touch in which mechanical stimuli stretch hundreds to thousands of axon protrusions across an end organ, opening proximal, axonal Piezo2 channels and exciting the neuron.
Keywords: Piezo2; focused ion beam scanning electron microscopy; light touch; mechanosensory neurons; mechanotransduction; touch end organs.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests C.S.X. is the inventor of a US patent assigned to HHMI for the enhanced FIB-SEM systems used in this work: Xu, C.S., Hayworth, K.J., and Hess, H.F. (2020) Enhanced FIB-SEM systems for large-volume 3D imaging. US Patent 10,600,615, 24 Mar. 2020.
Figures
Update of
-
Three-dimensional reconstructions of mechanosensory end organs suggest a unifying mechanism underlying dynamic, light touch.bioRxiv [Preprint]. 2023 Mar 18:2023.03.17.533188. doi: 10.1101/2023.03.17.533188. bioRxiv. 2023. Update in: Neuron. 2023 Oct 18;111(20):3211-3229.e9. doi: 10.1016/j.neuron.2023.08.023. PMID: 36993253 Free PMC article. Updated. Preprint.
Comment in
-
Engineering mechanoreceptor feature selectivity.Neuron. 2023 Oct 18;111(20):3137-3139. doi: 10.1016/j.neuron.2023.09.033. Neuron. 2023. PMID: 37857088
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
