

A new twist on PIFE: photoisomerisation-related fluorescence enhancement
- PMID: 37726007
- PMCID: PMC10570931
- DOI: 10.1088/2050-6120/acfb58
A new twist on PIFE: photoisomerisation-related fluorescence enhancement
Abstract
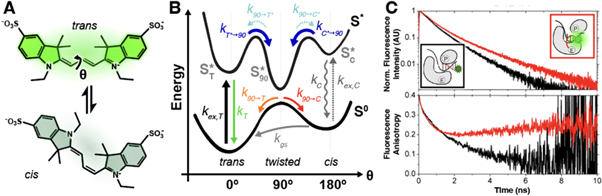
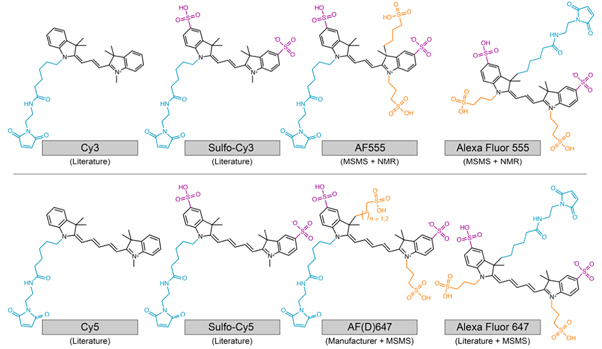
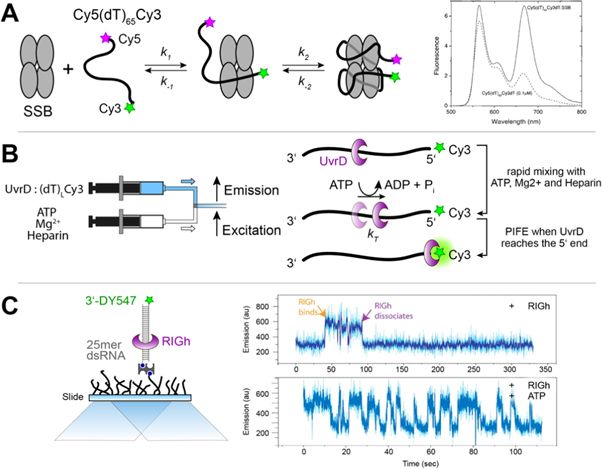
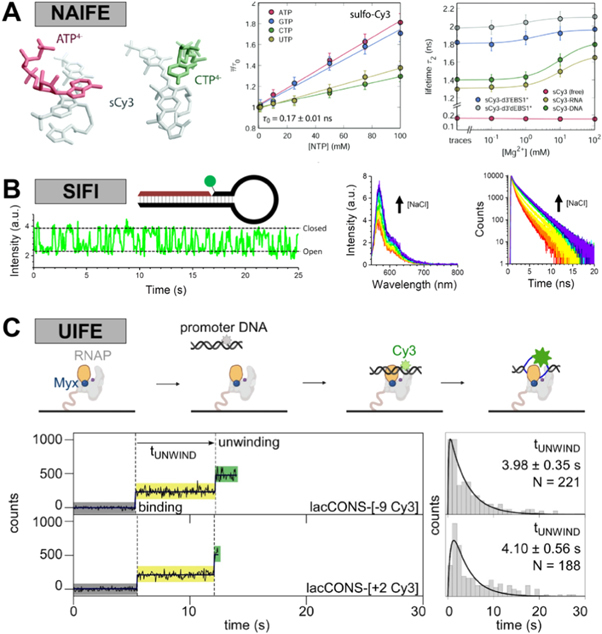
PIFE was first used as an acronym for protein-induced fluorescence enhancement, which refers to the increase in fluorescence observed upon the interaction of a fluorophore, such as a cyanine, with a protein. This fluorescence enhancement is due to changes in the rate ofcis/transphotoisomerisation. It is clear now that this mechanism is generally applicable to interactions with any biomolecule. In this review, we propose that PIFE is thereby renamed according to its fundamental working principle as photoisomerisation-related fluorescence enhancement, keeping the PIFE acronym intact. We discuss the photochemistry of cyanine fluorophores, the mechanism of PIFE, its advantages and limitations, and recent approaches to turning PIFE into a quantitative assay. We provide an overview of its current applications to different biomolecules and discuss potential future uses, including the study of protein-protein interactions, protein-ligand interactions and conformational changes in biomolecules.
Keywords: PIFE; biophysics; fluorescence spectroscopy; photoisomerisation; single-molecule.
Creative Commons Attribution license.
Figures
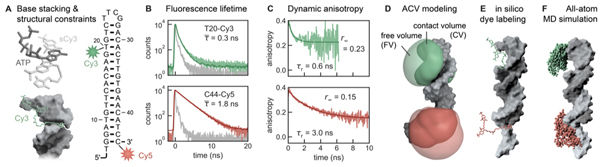
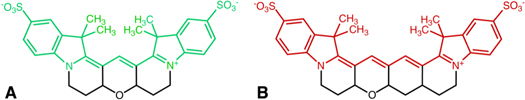
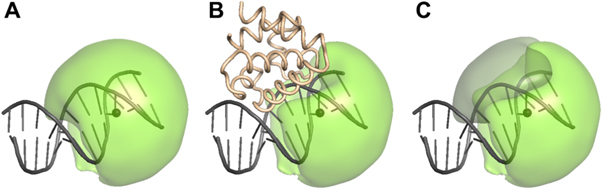
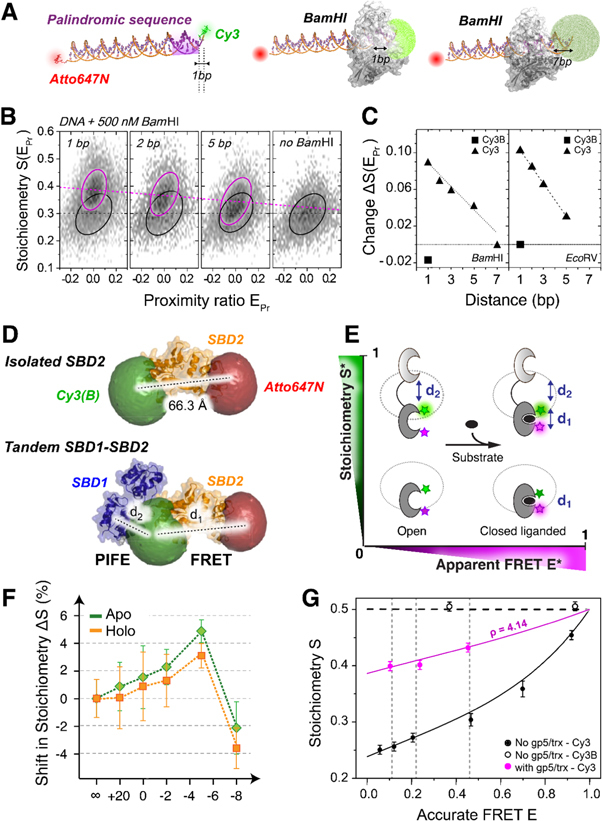
Update of
-
A new twist on PIFE: photoisomerisation-related fluorescence enhancement.ArXiv [Preprint]. 2023 Jul 10:arXiv:2302.12455v2. ArXiv. 2023. Update in: Methods Appl Fluoresc. 2023 Oct 12;12(1). doi: 10.1088/2050-6120/acfb58. PMID: 36866225 Free PMC article. Updated. Preprint.
References
-
- Lakowicz J R. Principles of Fluorescence Spectroscopy. New York, NY: Springer; 2006.
-
- Valeur B, Berberan-Santos M N. Molecular Fluorescence: Principles and Applications. Wiley; 2012.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources