

Epithelial plasticity and innate immune activation promote lung tissue remodeling following respiratory viral infection
- PMID: 37726288
- PMCID: PMC10509177
- DOI: 10.1038/s41467-023-41387-3
Epithelial plasticity and innate immune activation promote lung tissue remodeling following respiratory viral infection
Abstract
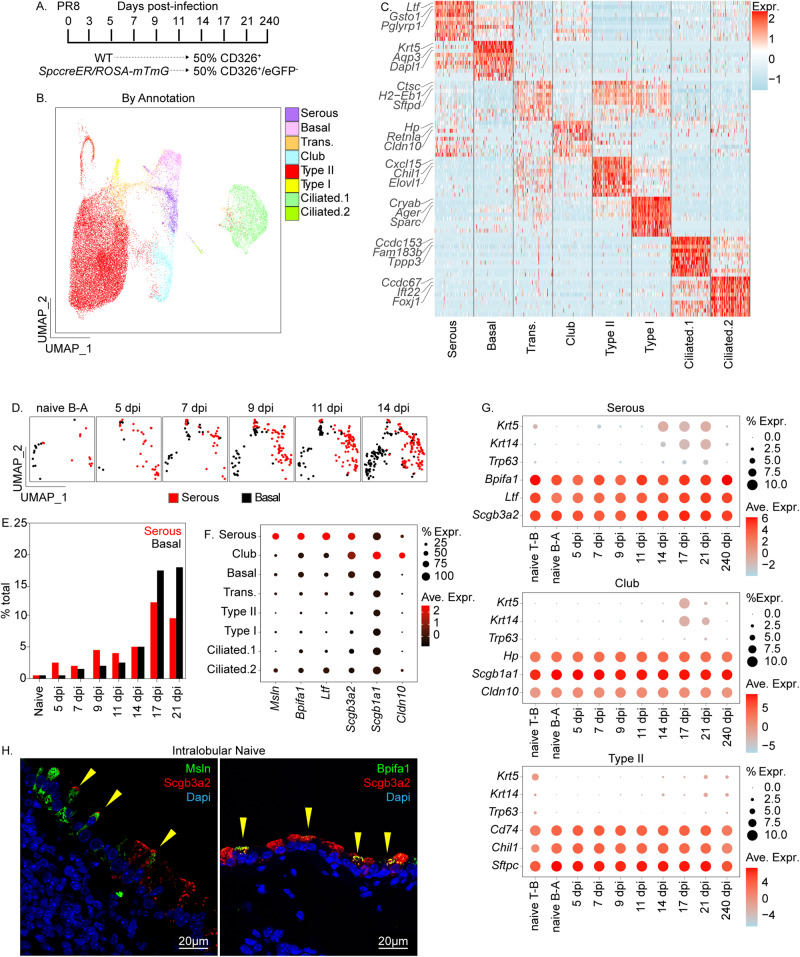
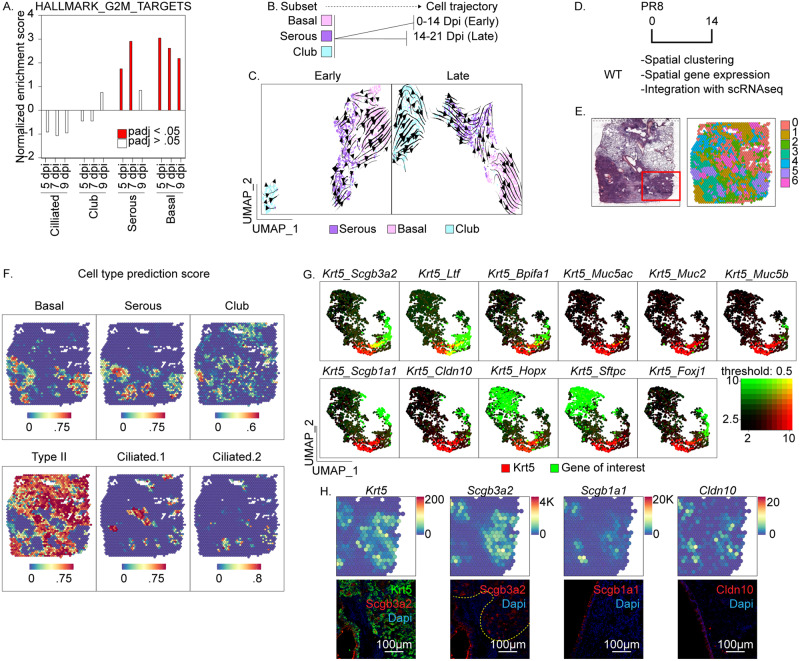
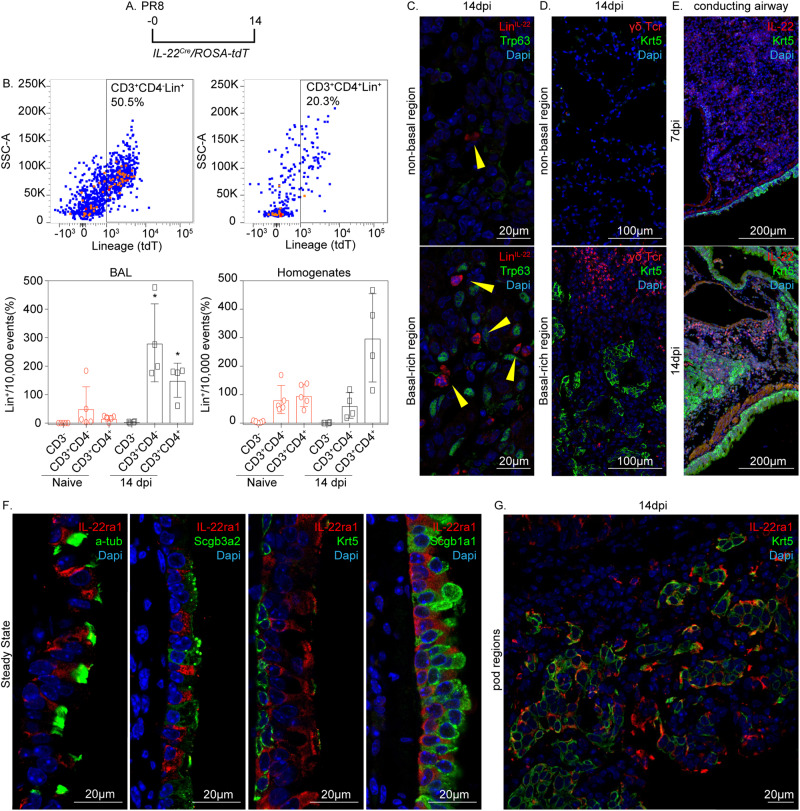
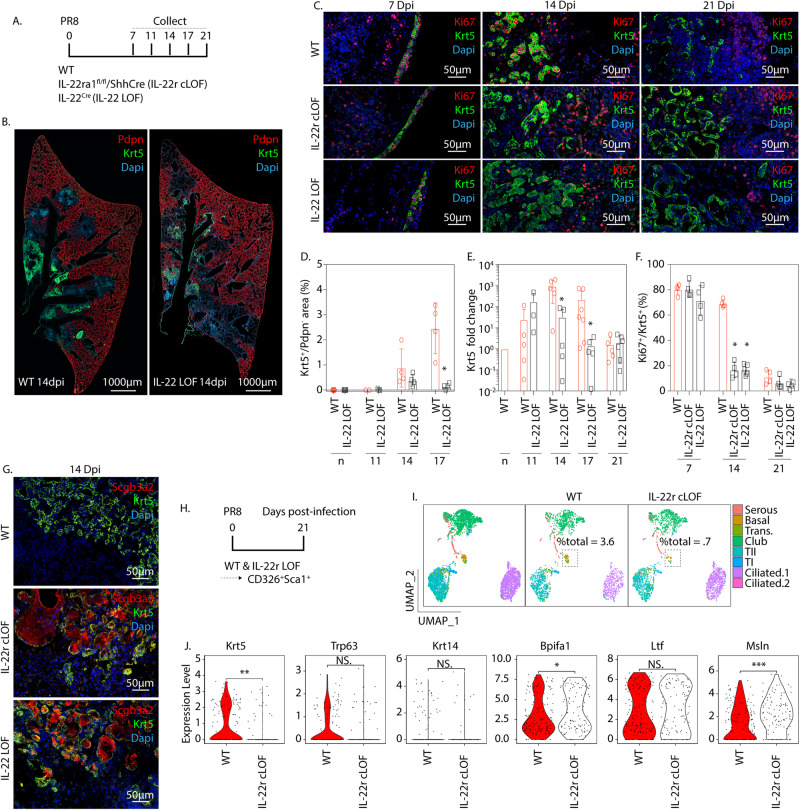
Epithelial plasticity has been suggested in lungs of mice following genetic depletion of stem cells but is of unknown physiological relevance. Viral infection and chronic lung disease share similar pathological features of stem cell loss in alveoli, basal cell (BC) hyperplasia in small airways, and innate immune activation, that contribute to epithelial remodeling and loss of lung function. We show that a subset of distal airway secretory cells, intralobar serous (IS) cells, are activated to assume BC fates following influenza virus infection. Injury-induced hyperplastic BC (hBC) differ from pre-existing BC by high expression of IL-22Ra1 and undergo IL-22-dependent expansion for colonization of injured alveoli. Resolution of virus-elicited inflammation results in BC to IS re-differentiation in repopulated alveoli, and increased local expression of protective antimicrobial factors, but fails to restore normal alveolar epithelium responsible for gas exchange.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
