

Efficient screening of adsorbed receptors for Salmonella phage LP31 and identification of receptor-binding protein
- PMID: 37728369
- PMCID: PMC10581130
- DOI: 10.1128/spectrum.02604-23
Efficient screening of adsorbed receptors for Salmonella phage LP31 and identification of receptor-binding protein
Abstract
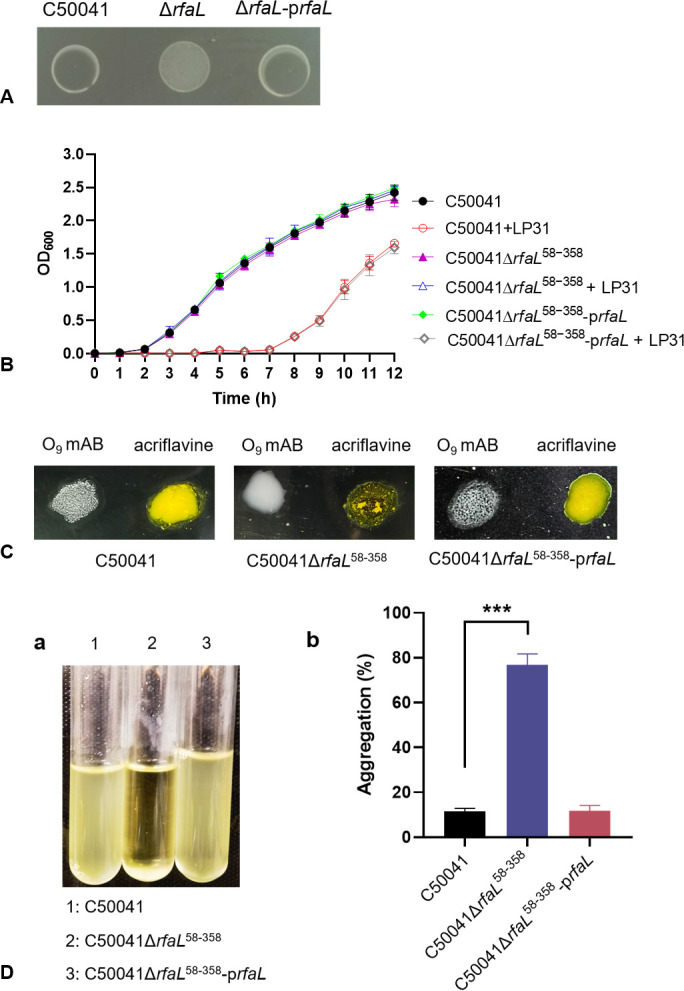
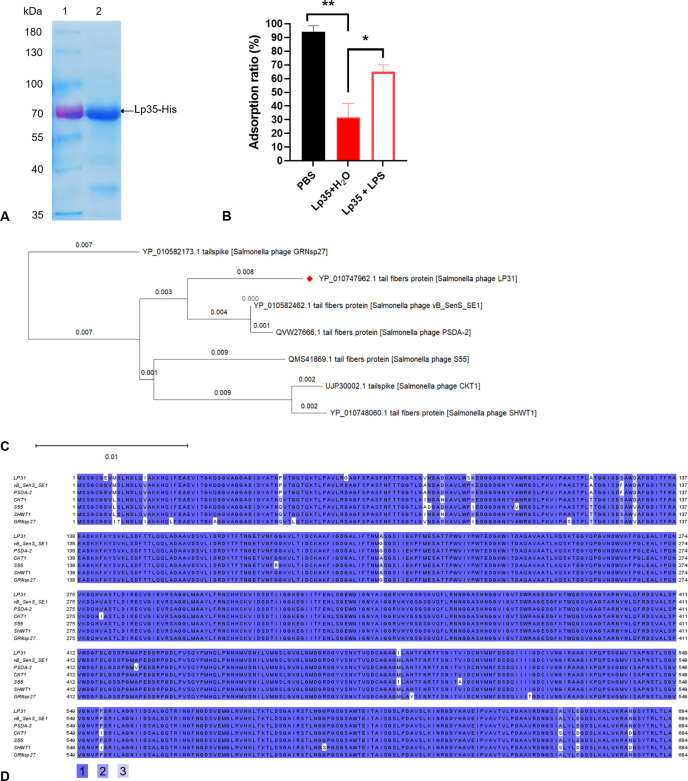
The adsorption process is the first step in the lifecycle of phages and plays a decisive role in the entire infection process. Identifying the adsorption mechanism of phages not only makes phage therapy more precise and efficient but also enables the exploration of other potential applications and modifications of phages. Phage LP31 can lyse multiple Salmonella serotypes, efficiently clearing biofilms formed by Salmonella enterica serovar Enteritidis (S. Enteritidis) and significantly reducing the concentration of S. Enteritidis in chicken feces. Therefore, LP31 has great potential for many practical applications. In this study, we established an efficient screening method for phage infection-related genes and identified a total of 10 genes related to the adsorption process of phage LP31. After the construction of strain C50041ΔrfaL 58-358, it was found that the knockout strain had a rough phenotype as an O-antigen-deficient strain. Adsorption rate and transmission electron microscopy experiments showed that the receptor for phage LP31 was the O9 antigen of S. Enteritidis. Homology comparison and adsorption experiments confirmed that the tail fiber protein Lp35 of phage LP31 participated in the adsorption process as a receptor-binding protein. IMPORTANCE A full understanding of the interaction between phages and their receptors can help with the development of phage-related products. Phages like LP31 with the tail fiber protein Lp35, or a closely related protein, have been reported to effectively recognize and infect multiple Salmonella serotypes. However, the role of these proteins in phage infection has not been previously described. In this study, we established an efficient screening method to detect phage adsorption to host receptors. We found that phage LP31 can utilize its tail fiber protein Lp35 to adsorb to the O9 antigen of S. Enteritidis, initiating the infection process. This study provides a great model system for further studies of how a phage-encoded receptor-binding protein (RBP) interacts with its host's RBP binding target, and this new model offers opportunities for further theoretical and experimental studies to understand the infection mechanism of phages.
Keywords: O9 antigen; Salmonella; adsorption receptor; phage; receptor-binding protein.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




Similar articles
-
A phage for the controlling of Salmonella in poultry and reducing biofilms.Vet Microbiol. 2022 Jun;269:109432. doi: 10.1016/j.vetmic.2022.109432. Epub 2022 Apr 14. Vet Microbiol. 2022. PMID: 35489296
-
Fitness Trade-Offs in Phage Cocktail-Resistant Salmonella enterica Serovar Enteritidis Results in Increased Antibiotic Susceptibility and Reduced Virulence.Microbiol Spectr. 2022 Oct 26;10(5):e0291422. doi: 10.1128/spectrum.02914-22. Epub 2022 Sep 27. Microbiol Spectr. 2022. PMID: 36165776 Free PMC article.
-
Identification of receptor-binding protein and host receptor of non-lytic dsRNA phage phiNY.Microbiol Spectr. 2024 Oct 22;12(12):e0146724. doi: 10.1128/spectrum.01467-24. Online ahead of print. Microbiol Spectr. 2024. PMID: 39436121 Free PMC article.
-
LamB, OmpC, and the Core Lipopolysaccharide of Escherichia coli K-12 Function as Receptors of Bacteriophage Bp7.J Virol. 2020 Jun 1;94(12):e00325-20. doi: 10.1128/JVI.00325-20. Print 2020 Jun 1. J Virol. 2020. PMID: 32238583 Free PMC article.
-
Phage Adsorption to Gram-Positive Bacteria.Viruses. 2023 Jan 10;15(1):196. doi: 10.3390/v15010196. Viruses. 2023. PMID: 36680236 Free PMC article. Review.
Cited by
-
PHIStruct: improving phage-host interaction prediction at low sequence similarity settings using structure-aware protein embeddings.Bioinformatics. 2024 Dec 26;41(1):btaf016. doi: 10.1093/bioinformatics/btaf016. Bioinformatics. 2024. PMID: 39804673 Free PMC article.
References
-
- Kosznik-Kwaśnicka K, Stasiłojć M, Grabowski Ł, Zdrojewska K, Węgrzyn G, Węgrzyn A. 2022. Efficacy and safety of phage therapy against Salmonella enterica serovars Typhimurium and Enteritidis estimated by using a battery of in vitro tests and the Galleria mellonella animal model. Microbiol Res 261:127052. doi:10.1016/j.micres.2022.127052 - DOI - PubMed
LinkOut - more resources
Full Text Sources
