

Prediction and design of protease enzyme specificity using a structure-aware graph convolutional network
- PMID: 37729196
- PMCID: PMC10523478
- DOI: 10.1073/pnas.2303590120
Prediction and design of protease enzyme specificity using a structure-aware graph convolutional network
Abstract
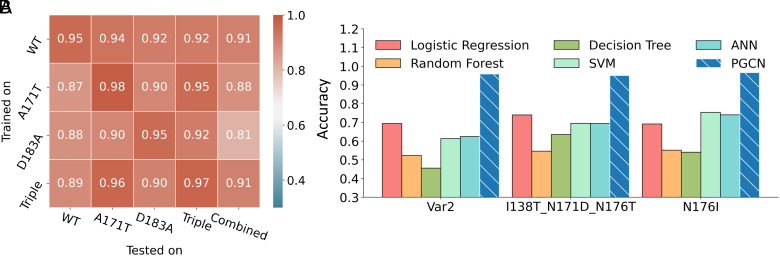
Site-specific proteolysis by the enzymatic cleavage of small linear sequence motifs is a key posttranslational modification involved in physiology and disease. The ability to robustly and rapidly predict protease-substrate specificity would also enable targeted proteolytic cleavage by designed proteases. Current methods for predicting protease specificity are limited to sequence pattern recognition in experimentally derived cleavage data obtained for libraries of potential substrates and generated separately for each protease variant. We reasoned that a more semantically rich and robust model of protease specificity could be developed by incorporating the energetics of molecular interactions between protease and substrates into machine learning workflows. We present Protein Graph Convolutional Network (PGCN), which develops a physically grounded, structure-based molecular interaction graph representation that describes molecular topology and interaction energetics to predict enzyme specificity. We show that PGCN accurately predicts the specificity landscapes of several variants of two model proteases. Node and edge ablation tests identified key graph elements for specificity prediction, some of which are consistent with known biochemical constraints for protease:substrate recognition. We used a pretrained PGCN model to guide the design of protease libraries for cleaving two noncanonical substrates, and found good agreement with experimental cleavage results. Importantly, the model can accurately assess designs featuring diversity at positions not present in the training data. The described methodology should enable the structure-based prediction of specificity landscapes of a wide variety of proteases and the construction of tailor-made protease editors for site-selectively and irreversibly modifying chosen target proteins.
Keywords: geometric machine learning; machine learning; protease specificity; protein design; yeast surface display.
Conflict of interest statement
The authors declare no competing interest.
Figures






Update of
-
Prediction and Design of Protease Enzyme Specificity Using a Structure-Aware Graph Convolutional Network.bioRxiv [Preprint]. 2023 Feb 16:2023.02.16.528728. doi: 10.1101/2023.02.16.528728. bioRxiv. 2023. Update in: Proc Natl Acad Sci U S A. 2023 Sep 26;120(39):e2303590120. doi: 10.1073/pnas.2303590120. PMID: 36824945 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
