

Olfactory bulb activity shapes the development of entorhinal-hippocampal coupling and associated cognitive abilities
- PMID: 37729915
- PMCID: PMC10617757
- DOI: 10.1016/j.cub.2023.08.072
Olfactory bulb activity shapes the development of entorhinal-hippocampal coupling and associated cognitive abilities
Abstract
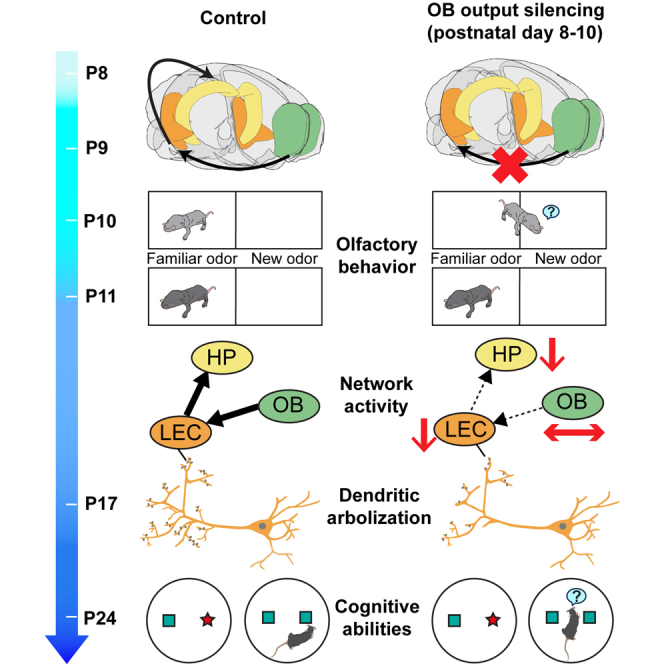
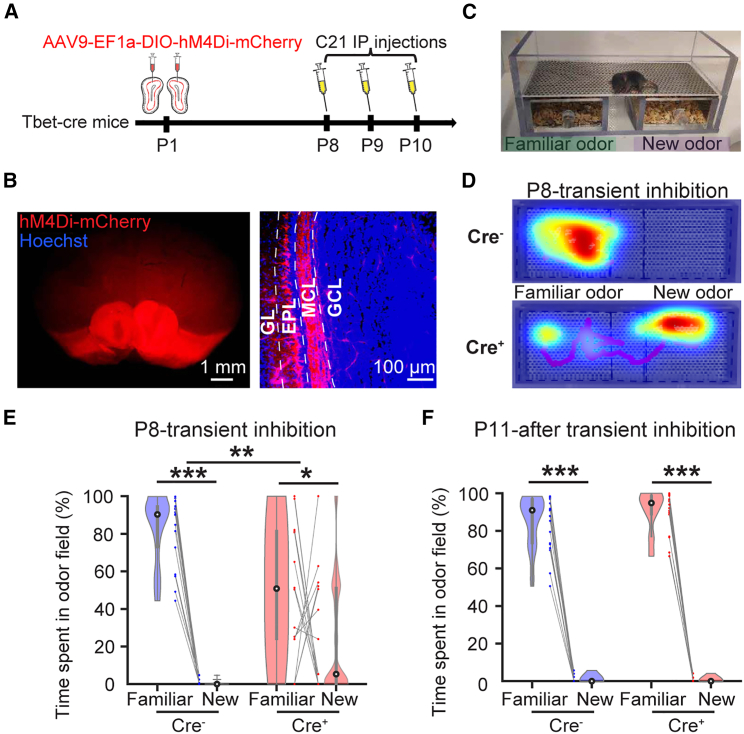
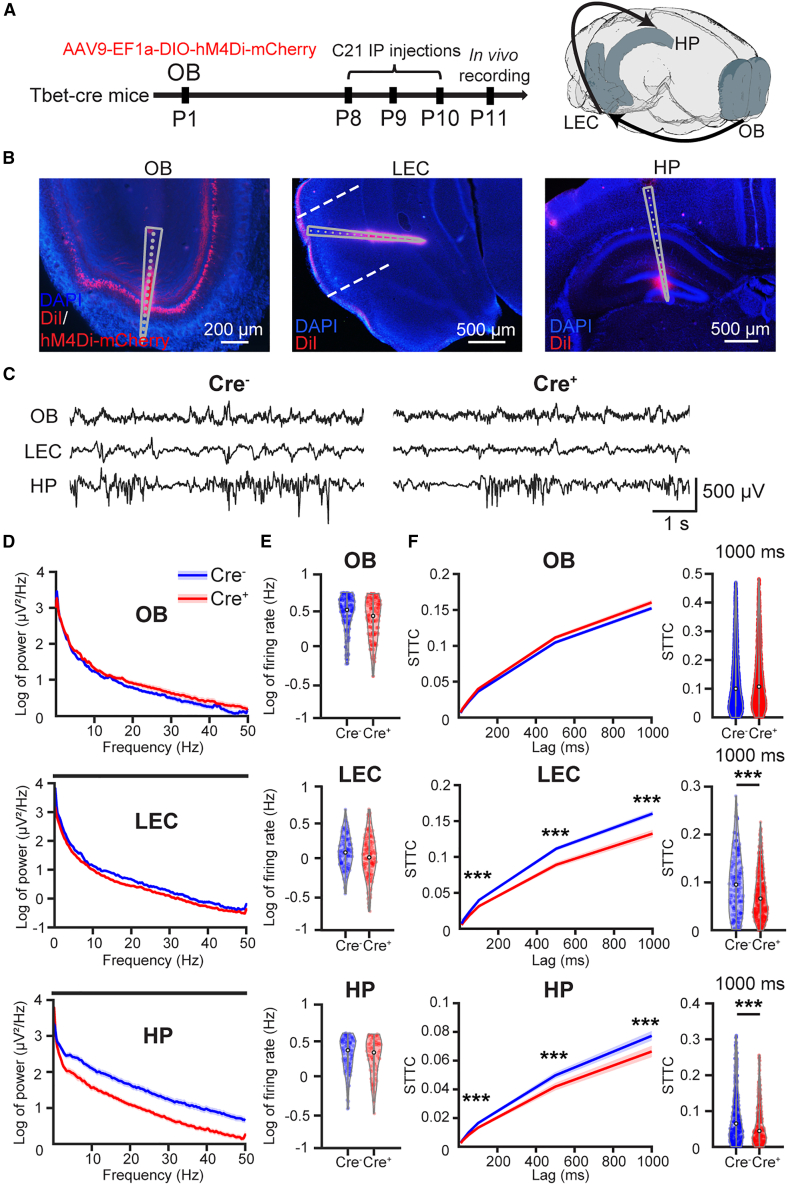
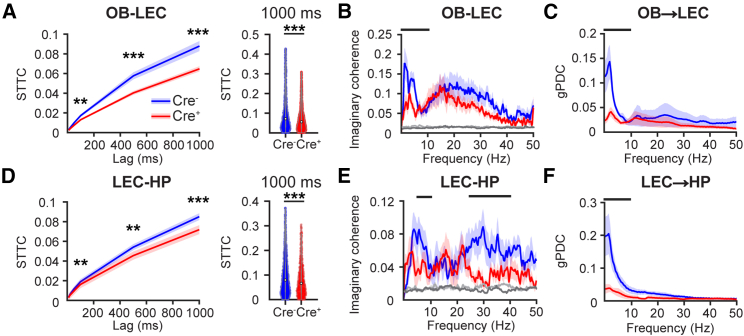
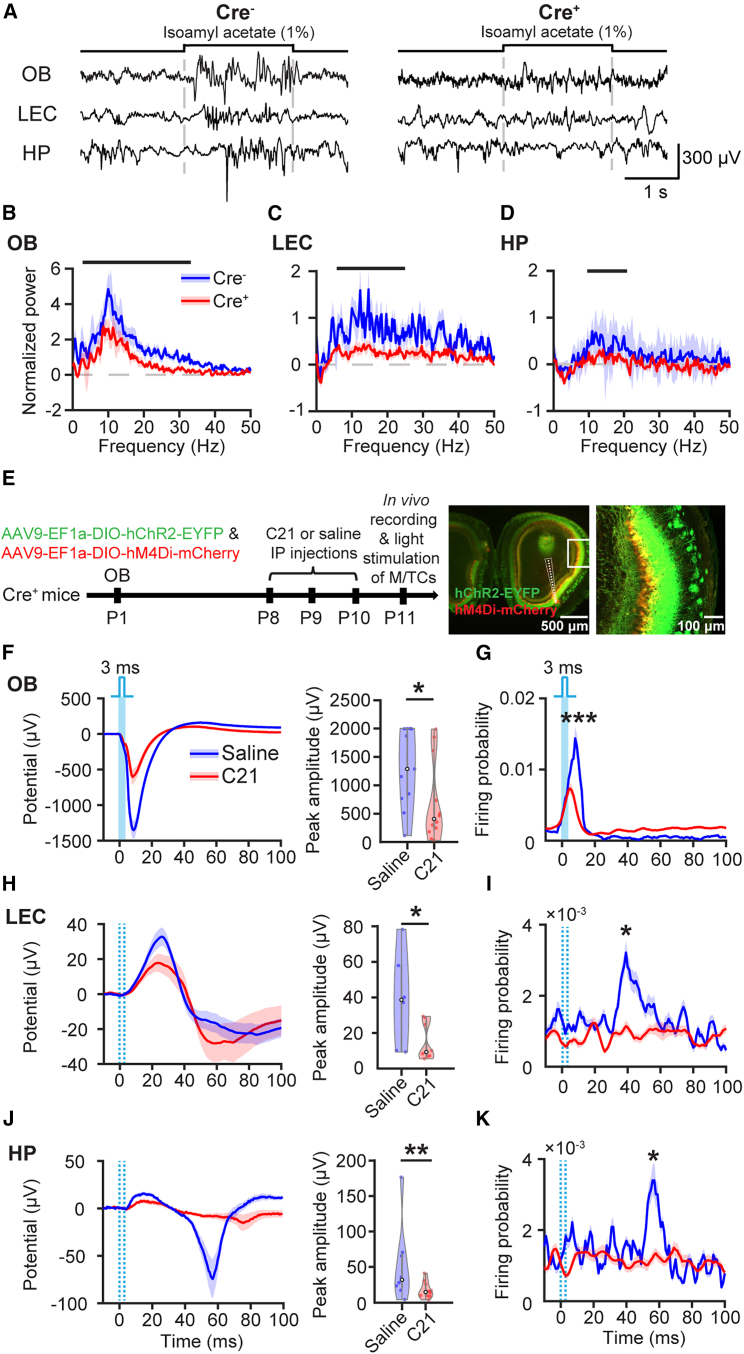
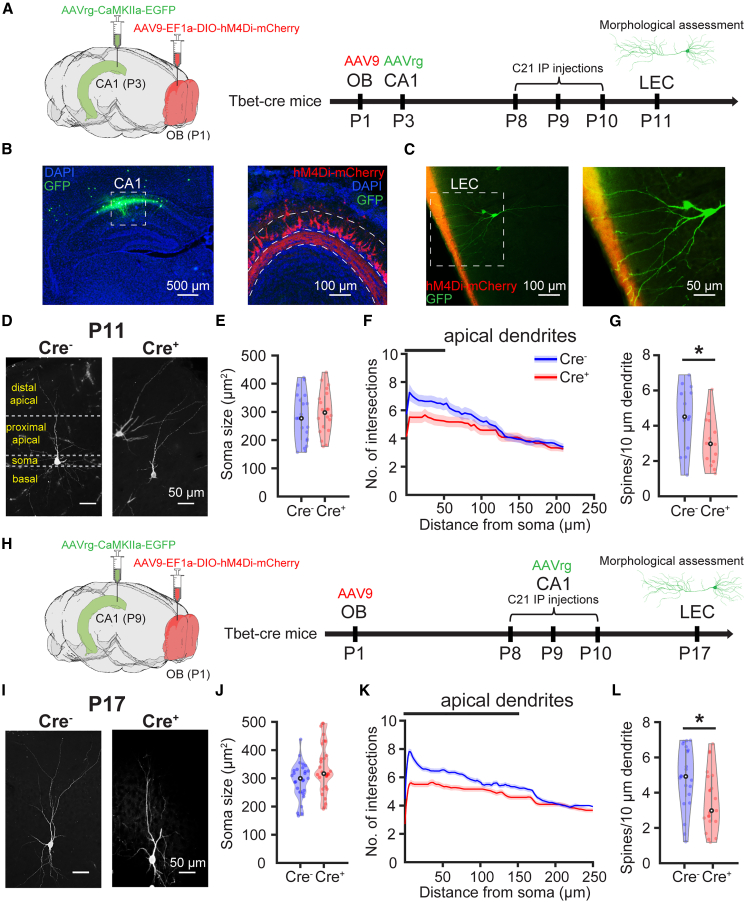
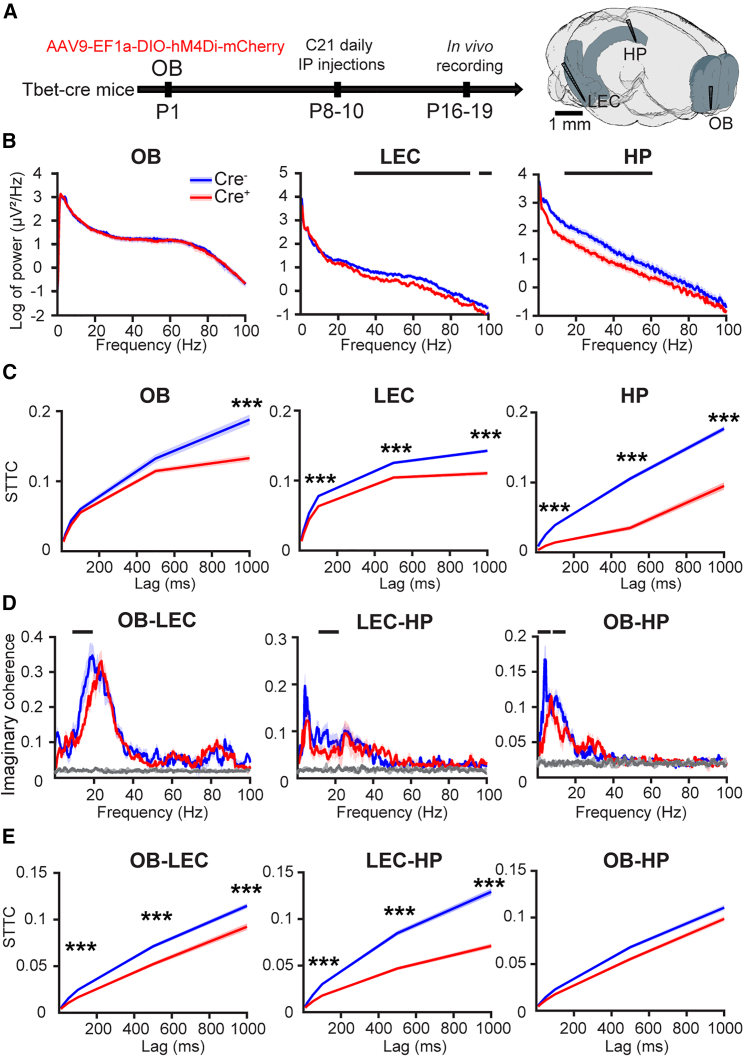
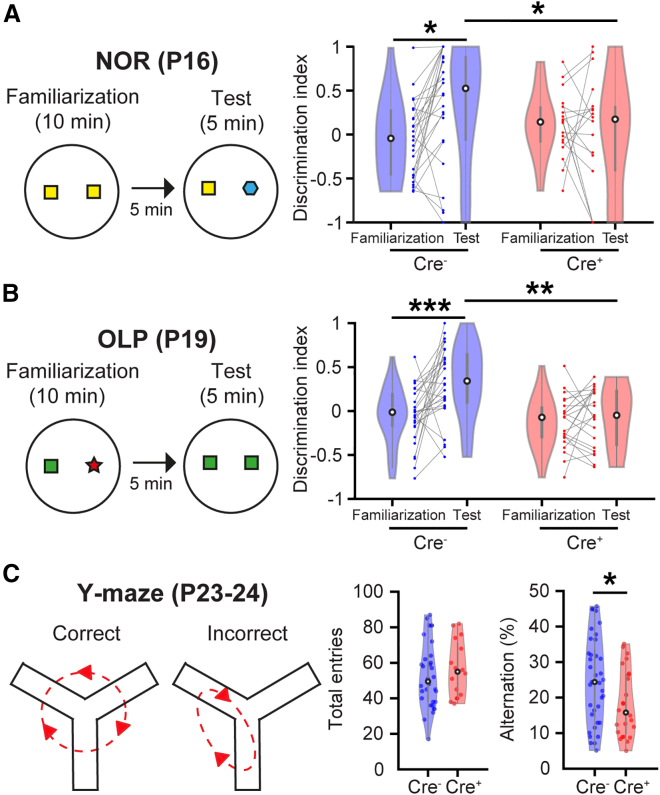
The interplay between olfaction and higher cognitive processing has been documented in the adult brain; however, its development is poorly understood. In mice, shortly after birth, endogenous and stimulus-evoked activity in the olfactory bulb (OB) boosts the oscillatory entrainment of downstream lateral entorhinal cortex (LEC) and hippocampus (HP). However, it is unclear whether early OB activity has a long-lasting impact on entorhinal-hippocampal function and cognitive processing. Here, we chemogenetically silenced the synaptic outputs of mitral/tufted cells, the main projection neurons in the OB, during postnatal days 8-10. The transient manipulation leads to a long-lasting reduction of oscillatory coupling and weaker responsiveness to stimuli within developing entorhinal-hippocampal circuits accompanied by dendritic sparsification of LEC pyramidal neurons. Moreover, the transient silencing reduces the performance in behavioral tests involving entorhinal-hippocampal circuits later in life. Thus, neonatal OB activity is critical for the functional LEC-HP development and maturation of cognitive abilities.
Keywords: chemogenetics; development; entorhinal-hippocampal network; olfactory bulb; oscillatory activity; recognition memory.
Copyright © 2023 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Neuroscience: Building better cognition through smell.Curr Biol. 2023 Oct 23;33(20):R1049-R1051. doi: 10.1016/j.cub.2023.09.030. Curr Biol. 2023. PMID: 37875078
References
-
- Quirk C.R., Zutshi I., Srikanth S., Fu M.L., Devico Marciano N., Wright M.K., Parsey D.F., Liu S., Siretskiy R.E., Huynh T.L., et al. Precisely timed theta oscillations are selectively required during the encoding phase of memory. Nat. Neurosci. 2021;24:1614–1627. doi: 10.1038/s41593-021-00919-0. - DOI - PMC - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
