

Genetic features and genomic targets of human KRAB-zinc finger proteins
- PMID: 37730438
- PMCID: PMC10547255
- DOI: 10.1101/gr.277722.123
Genetic features and genomic targets of human KRAB-zinc finger proteins
Abstract
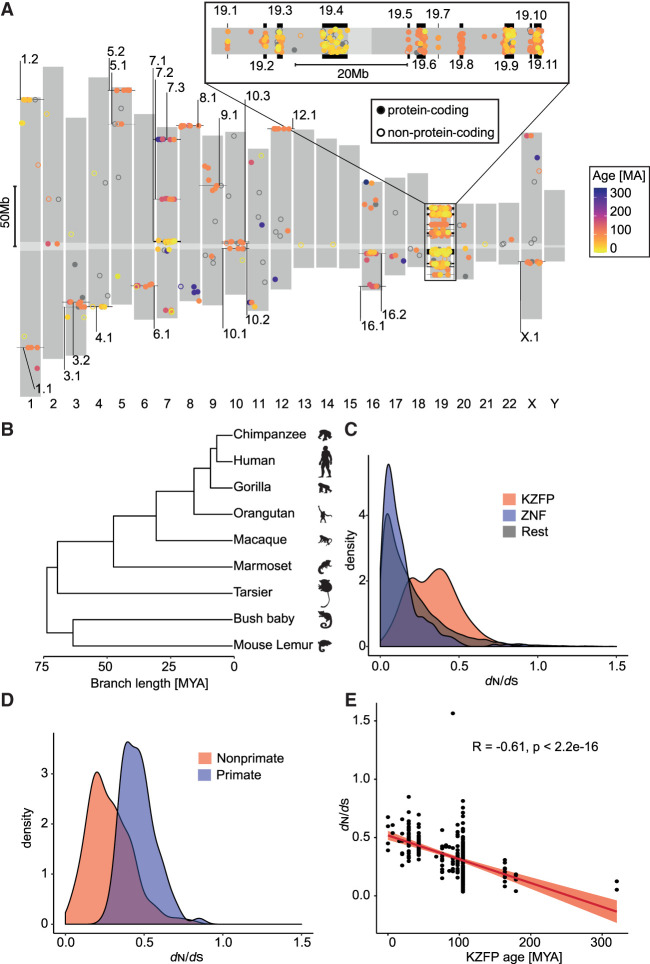
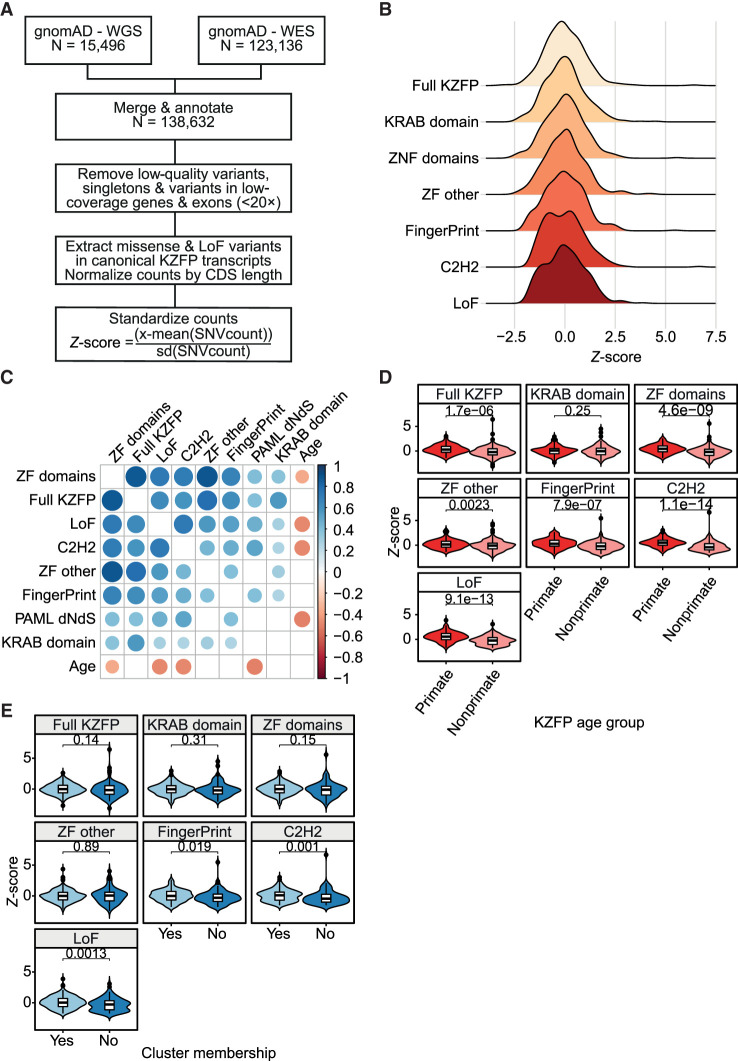
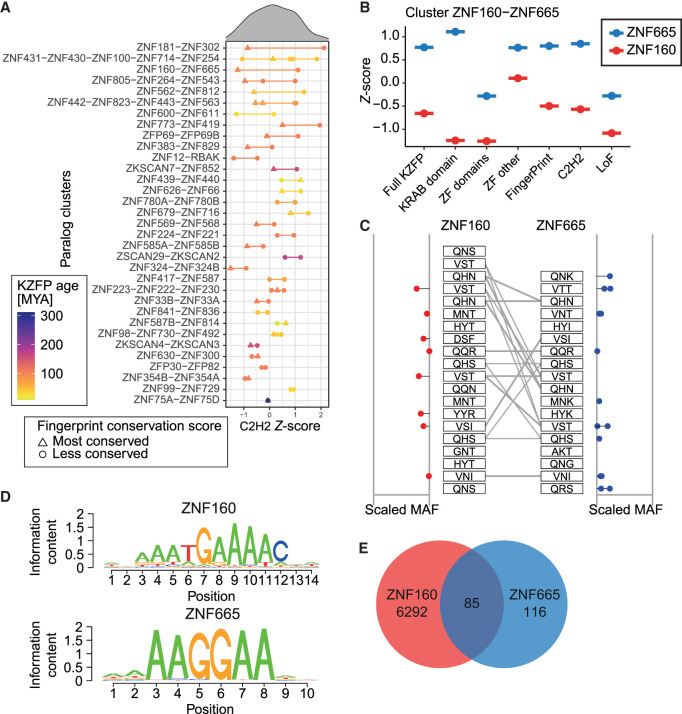
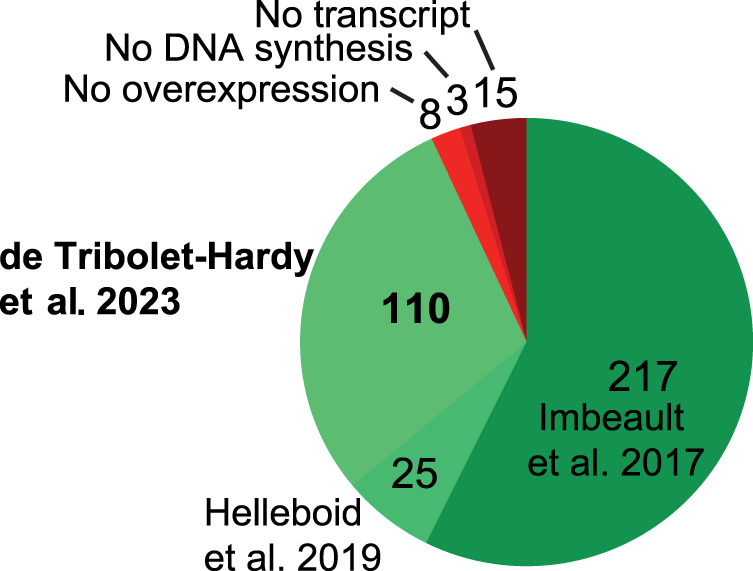
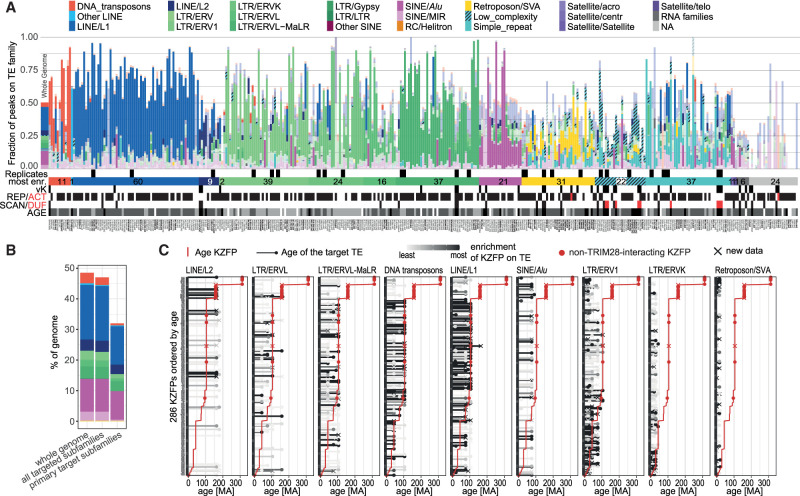
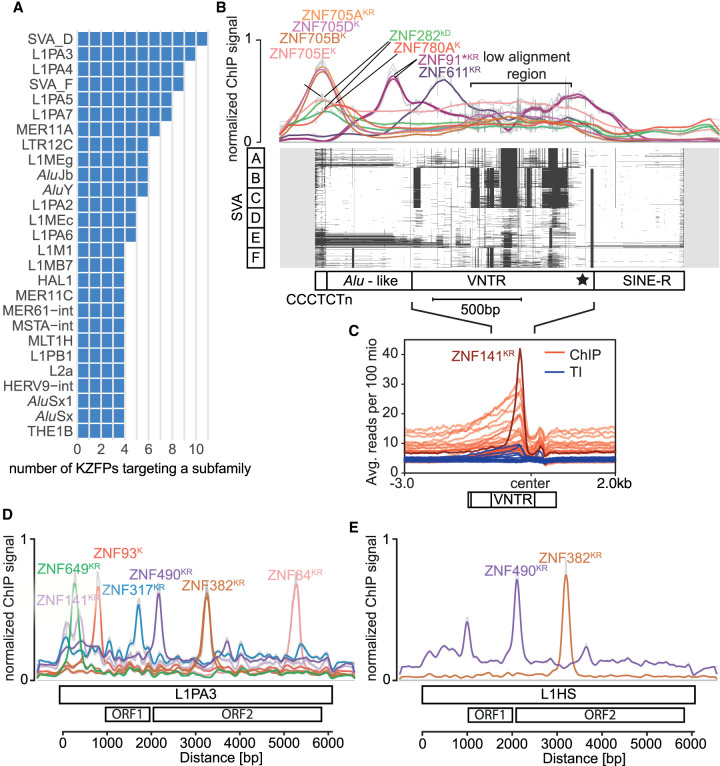
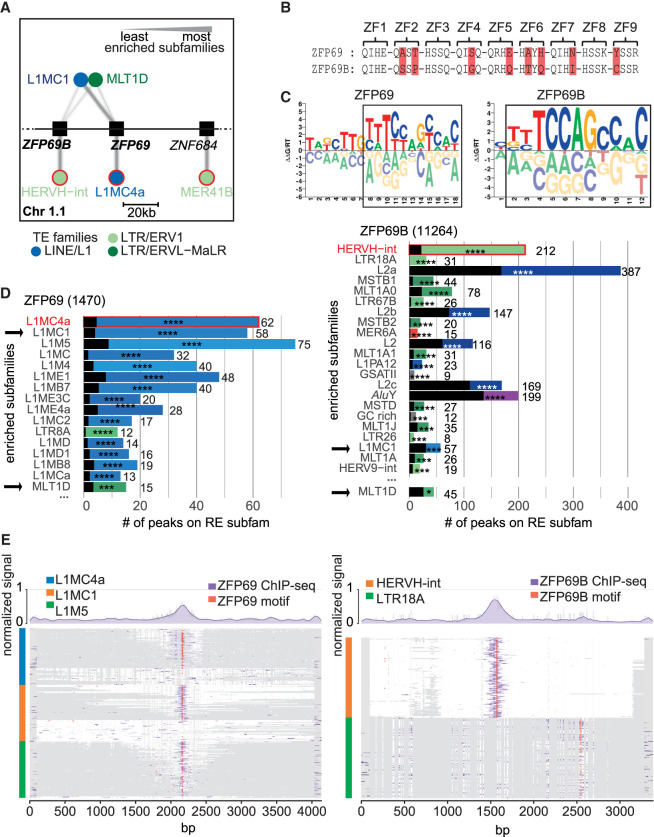
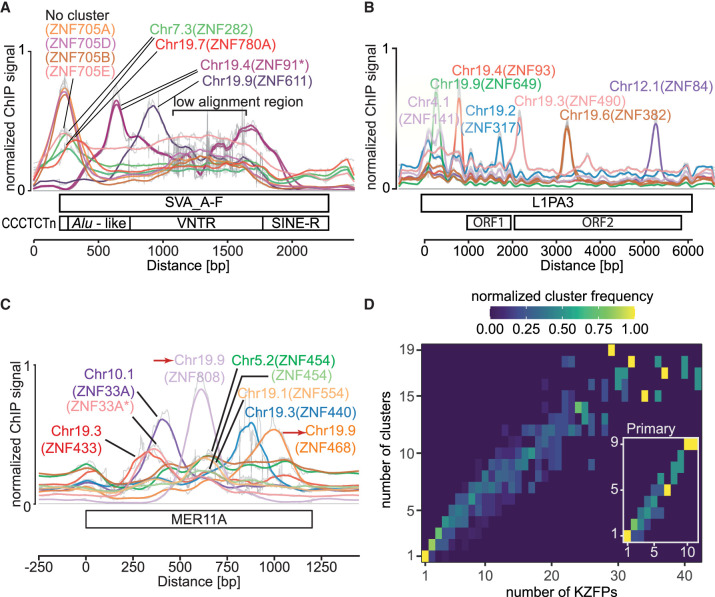
Krüppel-associated box (KRAB) domain-containing zinc finger proteins (KZFPs) are one of the largest groups of transcription factors encoded by tetrapods, with 378 members in human alone. KZFP genes are often grouped in clusters reflecting amplification by gene and segment duplication since the gene family first emerged more than 400 million years ago. Previous work has revealed that many KZFPs recognize transposable element (TE)-embedded sequences as genomic targets, and that KZFPs facilitate the co-option of the regulatory potential of TEs for the benefit of the host. Here, we present a comprehensive survey of the genetic features and genomic targets of human KZFPs, notably completing past analyses by adding data on close to a hundred family members. General principles emerge from our study of the TE-KZFP regulatory system, which point to multipronged evolutionary mechanisms underlaid by highly complex and combinatorial modes of action with strong influences on human speciation.
© 2023 de Tribolet-Hardy et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
-
- Arnold C, Matthews LJ, Nunn CL. 2010. The 10kTrees website: a new online resource for primate phylogeny. Evol Anthropol Issues News Rev 19: 114–118. 10.1002/evan.20251 - DOI
-
- Barnada SM, Isopi A, Tejada-Martinez D, Goubert C, Patoori S, Pagliaroli L, Tracewell M, Trizzino M. 2022. Genomic features underlie the co-option of SVA transposons as cis-regulatory elements in human pluripotent stem cells. PLoS Genet 18: e1010225. 10.1371/journal.pgen.1010225 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous