

Synergism between CMG helicase and leading strand DNA polymerase at replication fork
- PMID: 37730685
- PMCID: PMC10511561
- DOI: 10.1038/s41467-023-41506-0
Synergism between CMG helicase and leading strand DNA polymerase at replication fork
Abstract
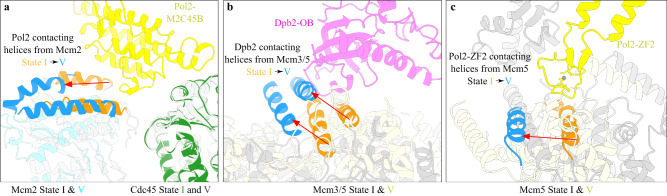
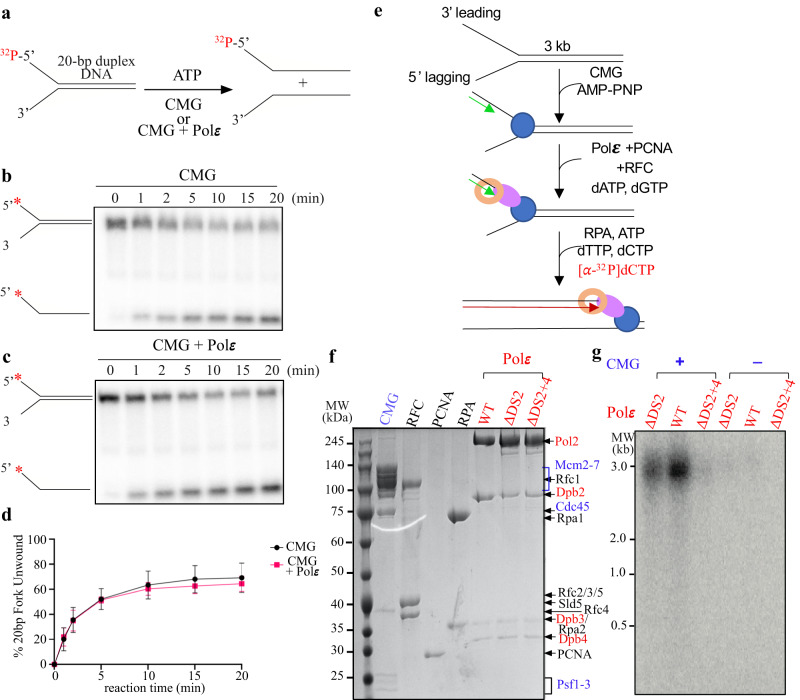
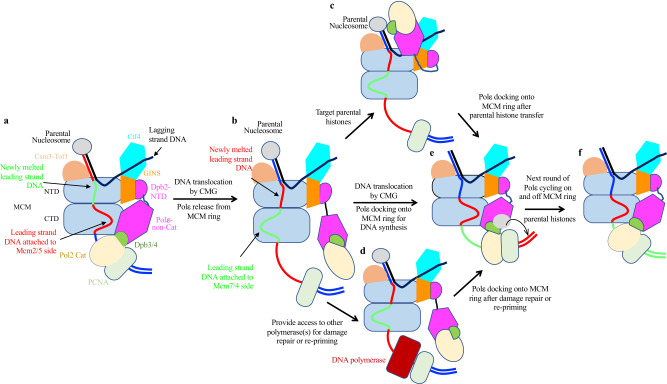
The replisome that replicates the eukaryotic genome consists of at least three engines: the Cdc45-MCM-GINS (CMG) helicase that separates duplex DNA at the replication fork and two DNA polymerases, one on each strand, that replicate the unwound DNA. Here, we determined a series of cryo-electron microscopy structures of a yeast replisome comprising CMG, leading-strand polymerase Polε and three accessory factors on a forked DNA. In these structures, Polε engages or disengages with the motor domains of the CMG by occupying two alternative positions, which closely correlate with the rotational movement of the single-stranded DNA around the MCM pore. During this process, the polymerase remains stably coupled to the helicase using Psf1 as a hinge. This synergism is modulated by a concerted rearrangement of ATPase sites to drive DNA translocation. The Polε-MCM coupling is not only required for CMG formation to initiate DNA replication but also facilitates the leading-strand DNA synthesis mediated by Polε. Our study elucidates a mechanism intrinsic to the replisome that coordinates the activities of CMG and Polε to negotiate any roadblocks, DNA damage, and epigenetic marks encountered during translocation along replication forks.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures
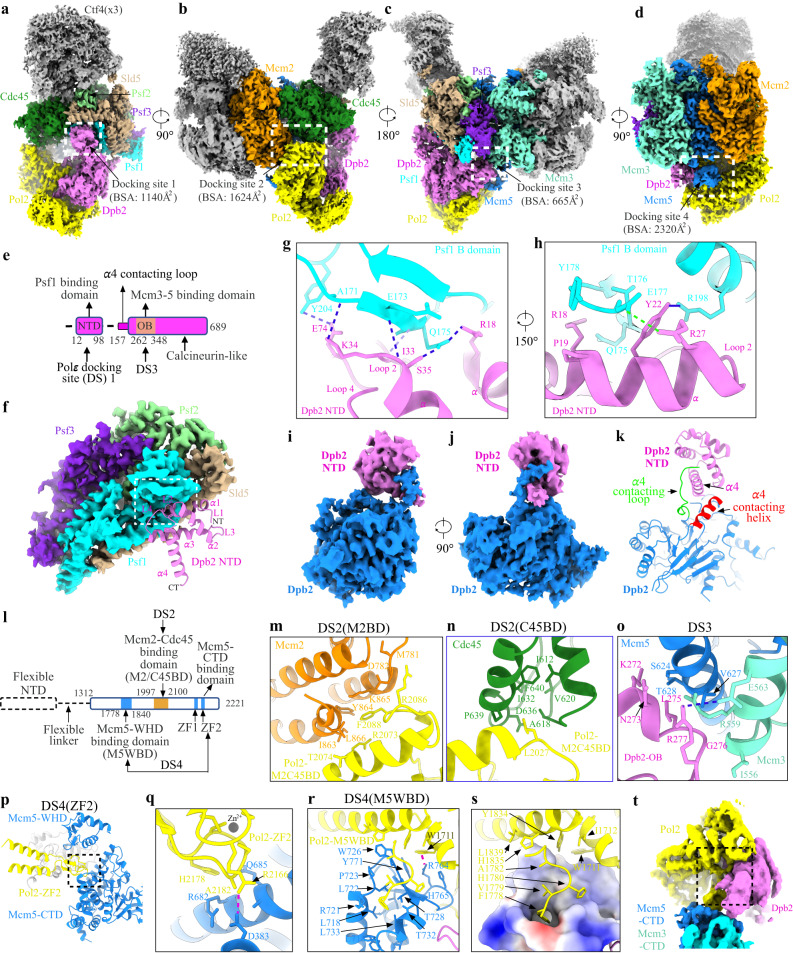
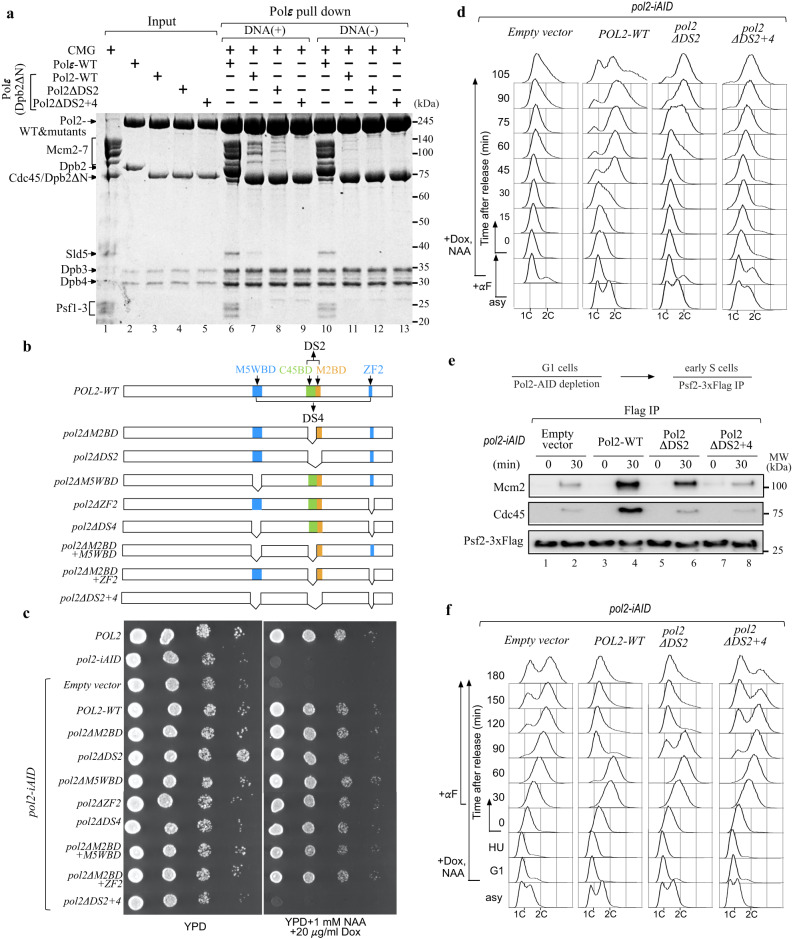
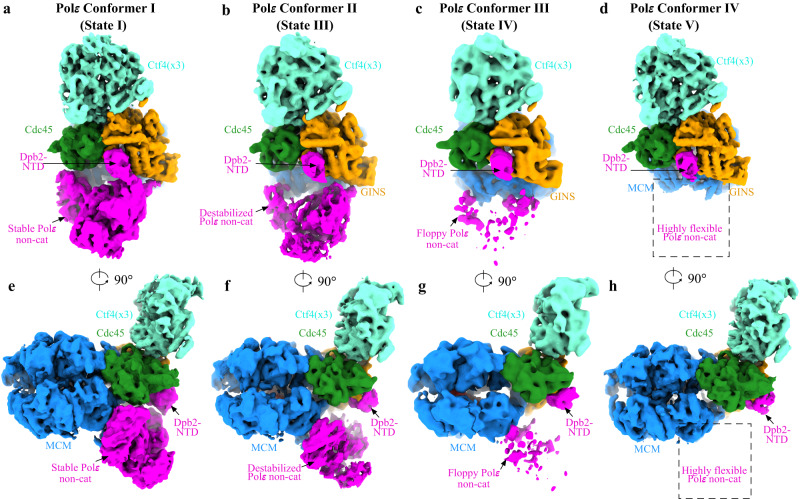
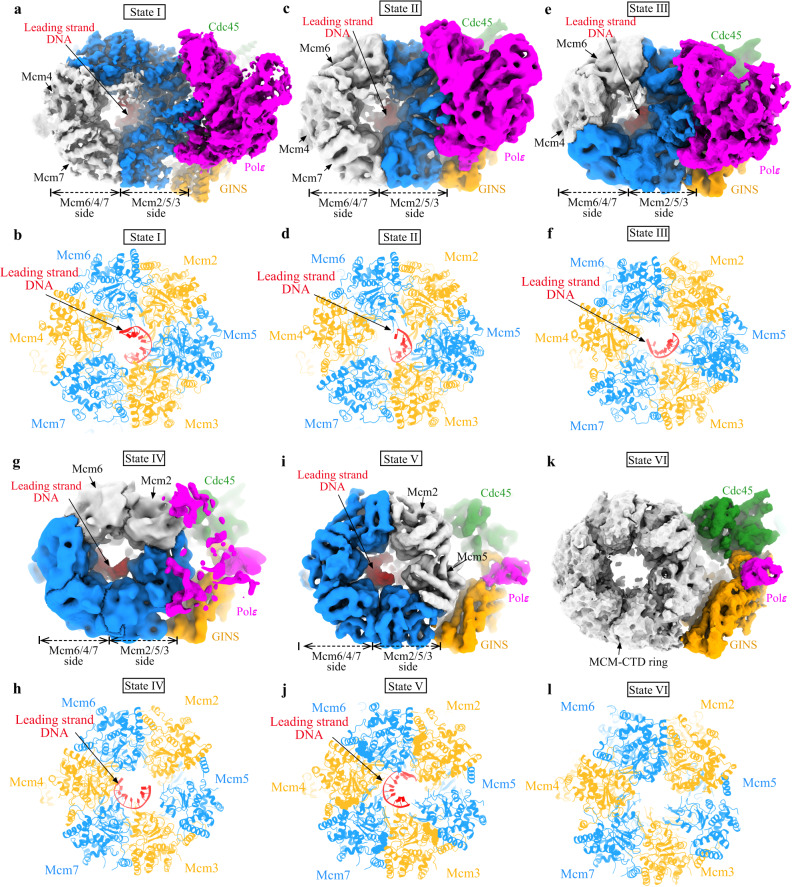
Similar articles
-
A common mechanism for recruiting the Rrm3 and RTEL1 accessory helicases to the eukaryotic replisome.EMBO J. 2024 Sep;43(18):3846-3875. doi: 10.1038/s44318-024-00168-4. Epub 2024 Jul 22. EMBO J. 2024. PMID: 39039288 Free PMC article.
-
Cryo-EM Structure of the Fork Protection Complex Bound to CMG at a Replication Fork.Mol Cell. 2020 Jun 4;78(5):926-940.e13. doi: 10.1016/j.molcel.2020.04.012. Epub 2020 May 4. Mol Cell. 2020. PMID: 32369734 Free PMC article.
-
Molecular Basis for ATP-Hydrolysis-Driven DNA Translocation by the CMG Helicase of the Eukaryotic Replisome.Cell Rep. 2019 Sep 3;28(10):2673-2688.e8. doi: 10.1016/j.celrep.2019.07.104. Cell Rep. 2019. PMID: 31484077 Free PMC article.
-
Arranging eukaryotic nuclear DNA polymerases for replication: Specific interactions with accessory proteins arrange Pols α, δ, and ϵ in the replisome for leading-strand and lagging-strand DNA replication.Bioessays. 2017 Aug;39(8):10.1002/bies.201700070. doi: 10.1002/bies.201700070. Bioessays. 2017. PMID: 28749073 Free PMC article. Review.
-
The eukaryotic CMG helicase pumpjack and integration into the replisome.Nucleus. 2016 Apr 25;7(2):146-54. doi: 10.1080/19491034.2016.1174800. Nucleus. 2016. PMID: 27310307 Free PMC article. Review.
Cited by
-
How similar are the molecular mechanisms of yeast and metazoan genome replication initiation?Biochem Soc Trans. 2025 Mar 7;53(2):353-61. doi: 10.1042/BST20220917. Biochem Soc Trans. 2025. PMID: 40052964 Free PMC article. Review.
-
Dynamic regulation of origin firing factors links CDK activity to dormant origin activation.bioRxiv [Preprint]. 2025 Jun 11:2025.06.10.657920. doi: 10.1101/2025.06.10.657920. bioRxiv. 2025. PMID: 40661636 Free PMC article. Preprint.
-
Parental histone transfer caught at the replication fork.Nature. 2024 Mar;627(8005):890-897. doi: 10.1038/s41586-024-07152-2. Epub 2024 Mar 6. Nature. 2024. PMID: 38448592
-
Starting DNA Synthesis: Initiation Processes during the Replication of Chromosomal DNA in Humans.Genes (Basel). 2024 Mar 14;15(3):360. doi: 10.3390/genes15030360. Genes (Basel). 2024. PMID: 38540419 Free PMC article. Review.
-
MCM5 UFMylation regulates replication origin firing and fork progression.EMBO J. 2025 Sep 12. doi: 10.1038/s44318-025-00562-6. Online ahead of print. EMBO J. 2025. PMID: 40940420
References
-
- Bleichert, F., Botchan, M. R. & Berger, J. M. Mechanisms for initiating cellular DNA replication. Science355, eaah6317 (2017). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
