

Cdk12 maintains the integrity of adult axons by suppressing actin remodeling
- PMID: 37730761
- PMCID: PMC10511712
- DOI: 10.1038/s41420-023-01642-4
Cdk12 maintains the integrity of adult axons by suppressing actin remodeling
Abstract
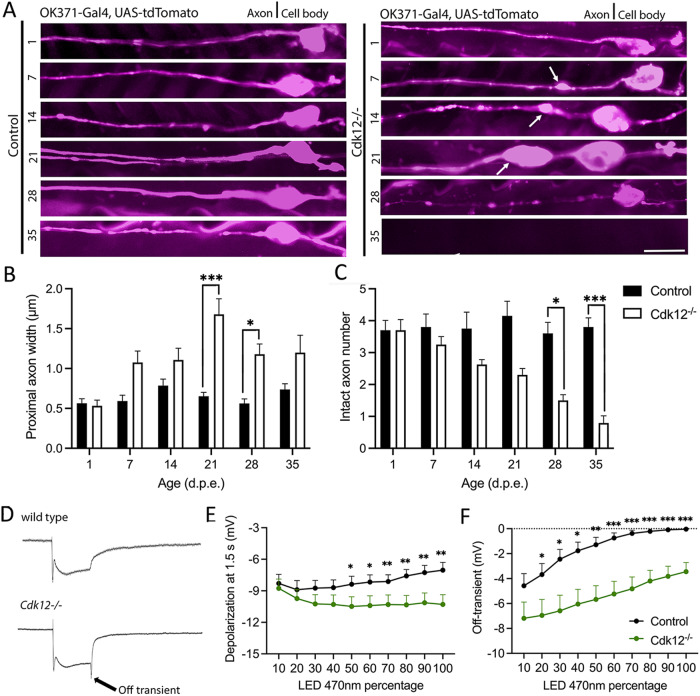
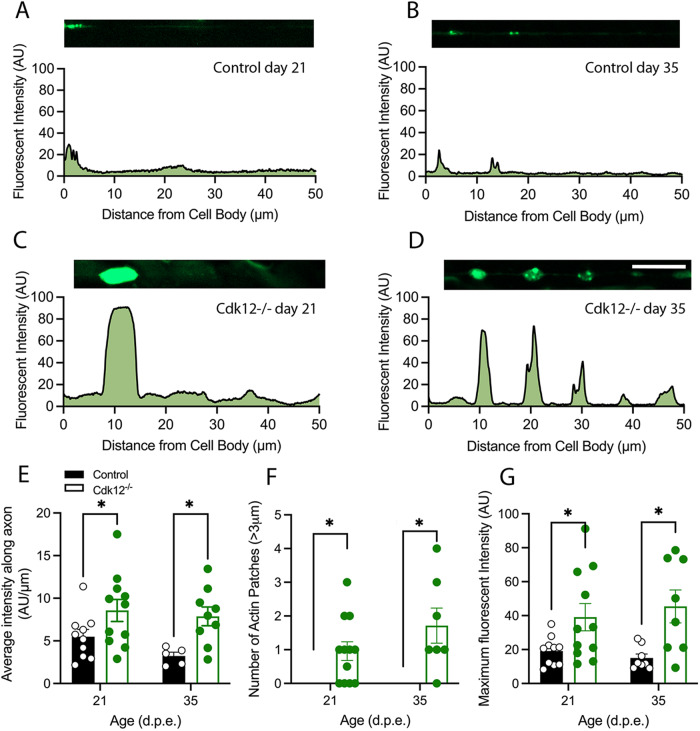
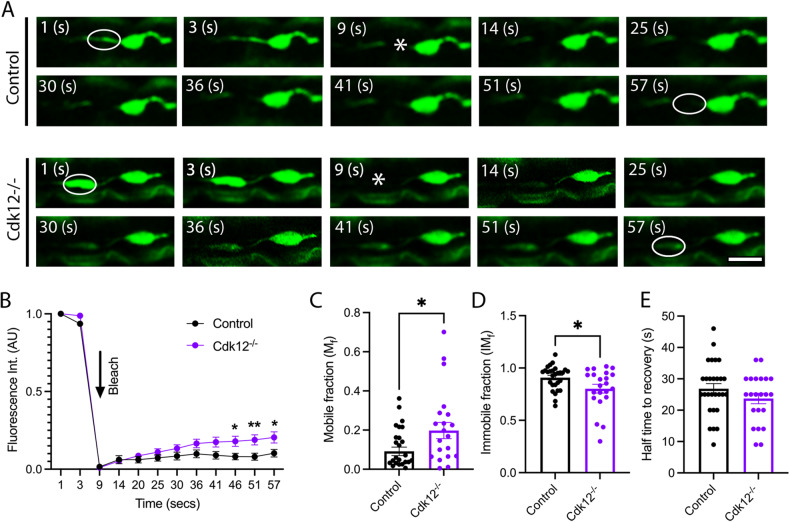
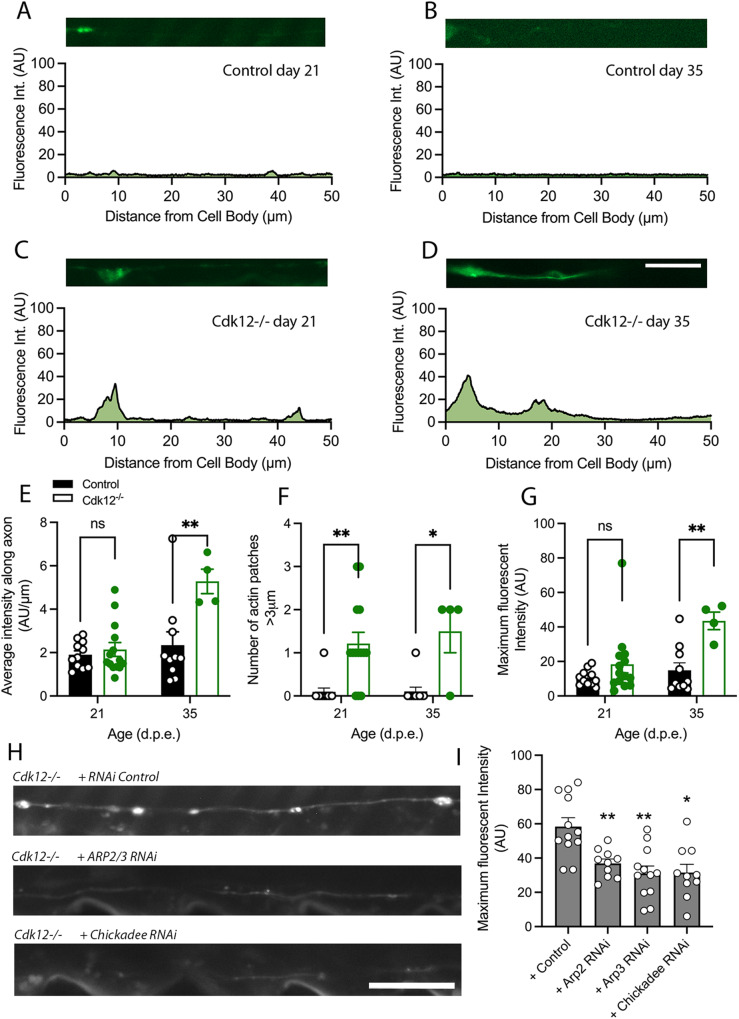
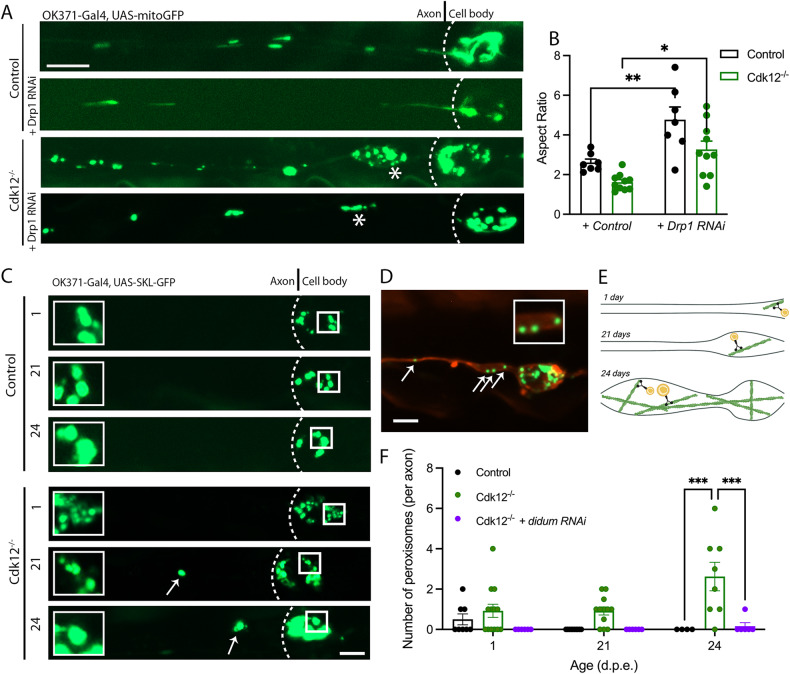
The role of cyclin-dependent kinases (CDKs) that are ubiquitously expressed in the adult nervous system remains unclear. Cdk12 is enriched in terminally differentiated neurons where its conical role in the cell cycle progression is redundant. We find that in adult neurons Cdk12 acts a negative regulator of actin formation, mitochondrial dynamics and neuronal physiology. Cdk12 maintains the size of the axon at sites proximal to the cell body through the transcription of homeostatic enzymes in the 1-carbon by folate pathway which utilize the amino acid homocysteine. Loss of Cdk12 leads to elevated homocysteine and in turn leads to uncontrolled F-actin formation and axonal swelling. Actin remodeling further induces Drp1-dependent fission of mitochondria and the breakdown of axon-soma filtration barrier allowing soma restricted cargos to enter the axon. We demonstrate that Cdk12 is also an essential gene for long-term neuronal survival and loss of this gene causes age-dependent neurodegeneration. Hyperhomocysteinemia, actin changes, and mitochondrial fragmentation are associated with several neurodegenerative conditions such as Alzheimer's disease and we provide a candidate molecular pathway to link together such pathological events.
© 2023. Cell Death Differentiation Association (ADMC).
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Cdk12 and Cdk13 regulate axonal elongation through a common signaling pathway that modulates Cdk5 expression.Exp Neurol. 2014 Nov;261:10-21. doi: 10.1016/j.expneurol.2014.06.024. Epub 2014 Jul 3. Exp Neurol. 2014. PMID: 24999027
-
Gene expression regulation by CDK12: a versatile kinase in cancer with functions beyond CTD phosphorylation.Exp Mol Med. 2020 May;52(5):762-771. doi: 10.1038/s12276-020-0442-9. Epub 2020 May 25. Exp Mol Med. 2020. PMID: 32451425 Free PMC article. Review.
-
Compartment-Specific Regulation of Autophagy in Primary Neurons.J Neurosci. 2016 Jun 1;36(22):5933-45. doi: 10.1523/JNEUROSCI.4401-15.2016. J Neurosci. 2016. PMID: 27251616 Free PMC article.
-
Actin(g) on mitochondria - a role for cofilin1 in neuronal cell death pathways.Biol Chem. 2019 Aug 27;400(9):1089-1097. doi: 10.1515/hsz-2019-0120. Biol Chem. 2019. PMID: 31256058 Review.
-
The emerging roles of CDK12 in tumorigenesis.Cell Div. 2017 Oct 27;12:7. doi: 10.1186/s13008-017-0033-x. eCollection 2017. Cell Div. 2017. PMID: 29090014 Free PMC article. Review.
Cited by
-
Cyclin-dependent kinases regulate the adult nervous system via the one-carbon-metabolism.Cell Death Dis. 2023 Jul 14;14(7):429. doi: 10.1038/s41419-023-05950-6. Cell Death Dis. 2023. PMID: 37452015 Free PMC article. No abstract available.
-
Physiological and pathological roles of the transcriptional kinases CDK12 and CDK13 in the central nervous system.Cell Death Differ. 2025 Mar;32(3):371-381. doi: 10.1038/s41418-024-01413-3. Epub 2024 Nov 12. Cell Death Differ. 2025. PMID: 39533070 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
