

Negative chemotaxis of Ligilactobacillus agilis BKN88 against gut-derived substances
- PMID: 37730901
- PMCID: PMC10511705
- DOI: 10.1038/s41598-023-42840-5
Negative chemotaxis of Ligilactobacillus agilis BKN88 against gut-derived substances
Abstract
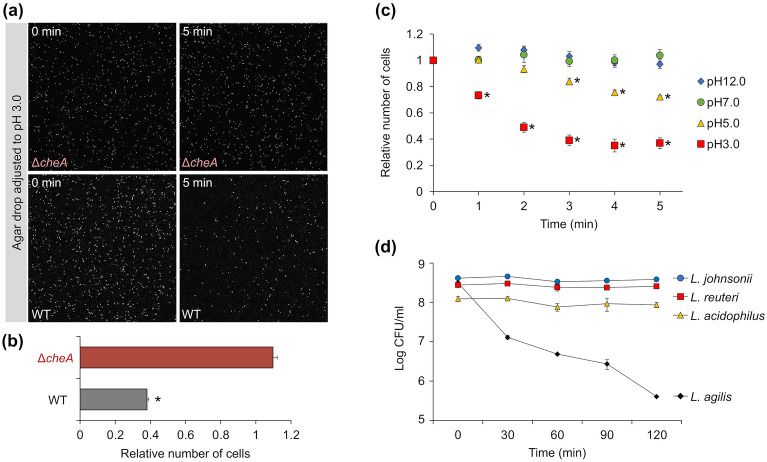
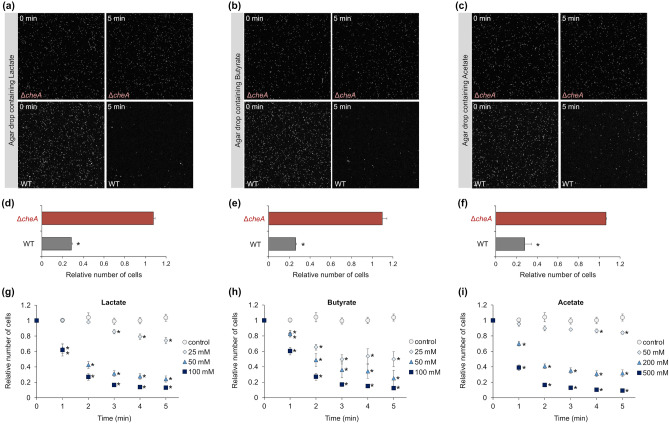
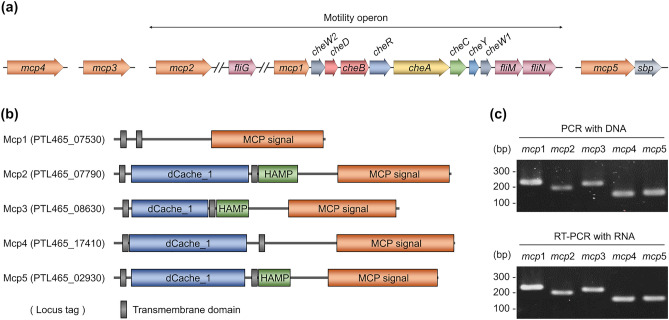
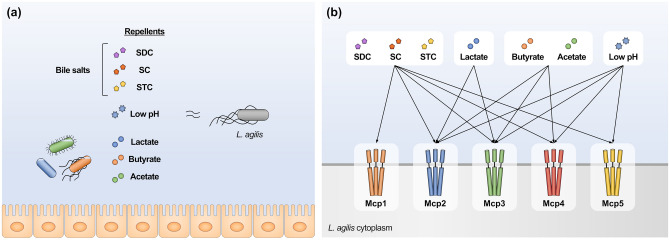
Ligilactobacillus agilis is a motile lactic acid bacterium found in the gastrointestinal tracts of animals. The findings of our previous study suggest that the motility of L. agilis BKN88 enables gut colonization in murine models. However, the chemotactic abilities of motile lactobacilli remain unknown. This study aimed to identify the gut-derived chemoeffectors and their corresponding chemoreceptors in L. agilis BKN88. Chemotaxis assays with chemotactic and non-chemotactic (ΔcheA) L. agilis strains revealed that low pH, organic acids, and bile salts served as repellents. L. agilis BKN88 was more sensitive to bile and acid than the gut-derived non-motile lactobacilli, implying that L. agilis might utilize motility and chemotaxis instead of exhibiting stress tolerance/resistance. L. agilis BKN88 contains five putative chemoreceptor genes (mcp1-mcp5). Chemotaxis assays using a series of chemoreceptor mutants revealed that each of the five chemoreceptors could sense multiple chemoeffectors and that these chemoreceptors were functionally redundant. Mcp2 and Mcp3 sensed all tested chemoeffectors. This study provides further insights into the interactions between chemoreceptors and ligands of motile lactobacilli and the unique ecological and evolutionary features of motile lactobacilli, which may be distinct from those of non-motile lactobacilli.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures


References
-
- Adler J. Chemotaxis in bacteria motile Escherichia coli migrate in bands that are influenced by oxygen and organic nutrients. Adv. Sci. 2008;153:708–716.
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
