

W546 stacking disruption traps the human porphyrin transporter ABCB6 in an outward-facing transient state
- PMID: 37735522
- PMCID: PMC10514269
- DOI: 10.1038/s42003-023-05339-3
W546 stacking disruption traps the human porphyrin transporter ABCB6 in an outward-facing transient state
Abstract
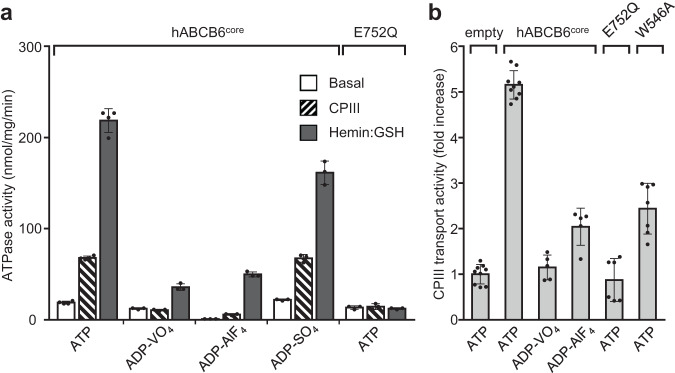
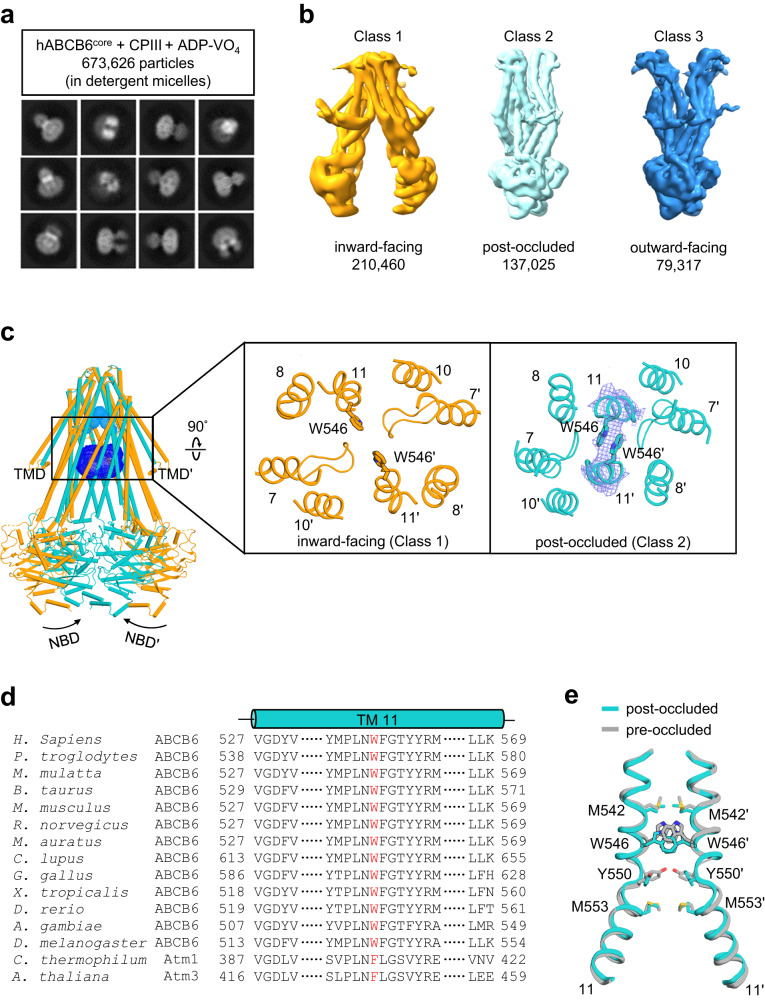
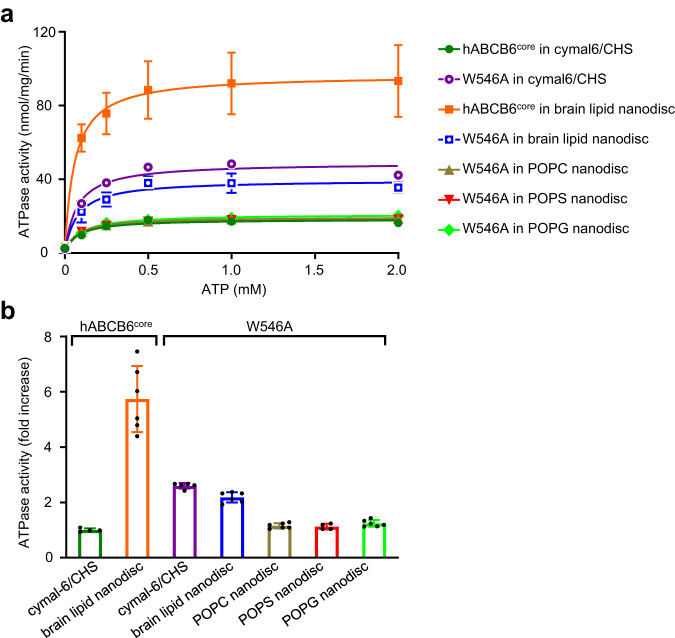
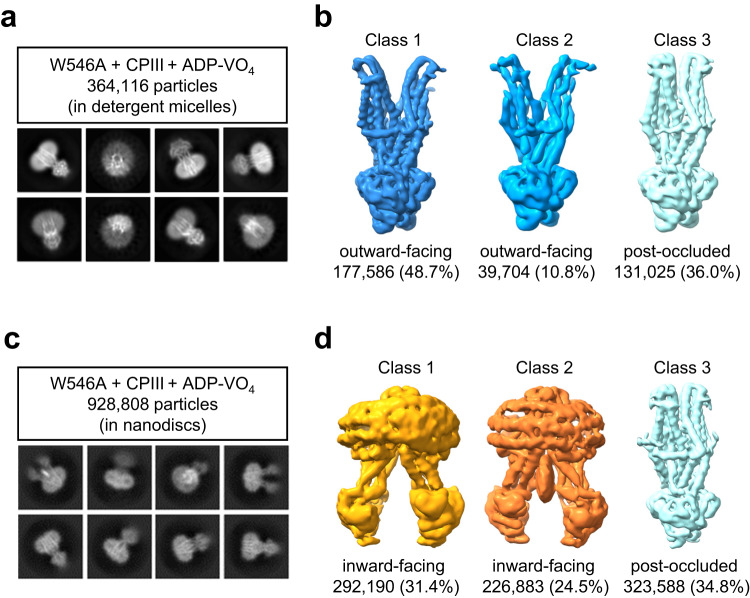
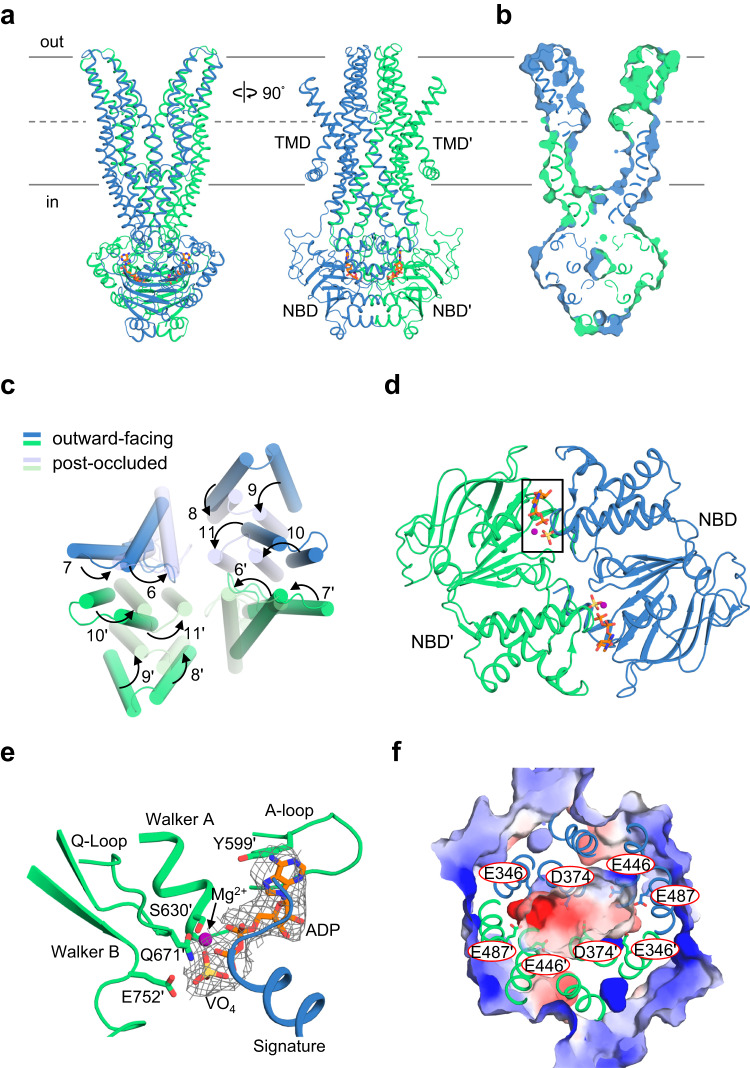
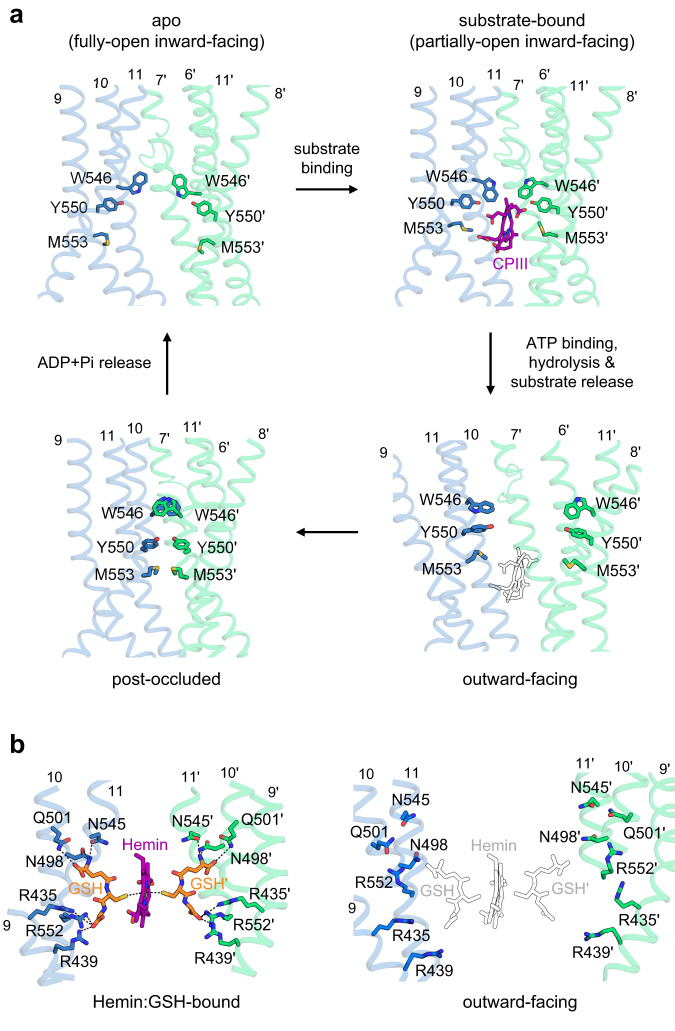
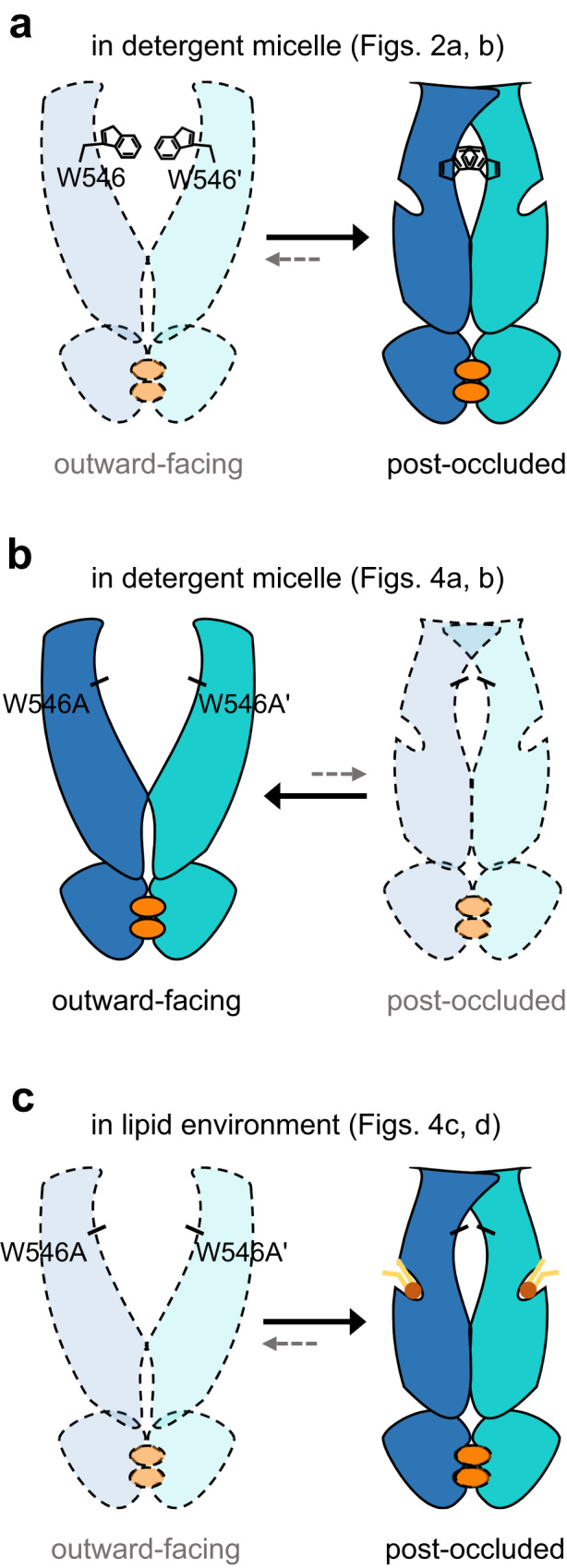
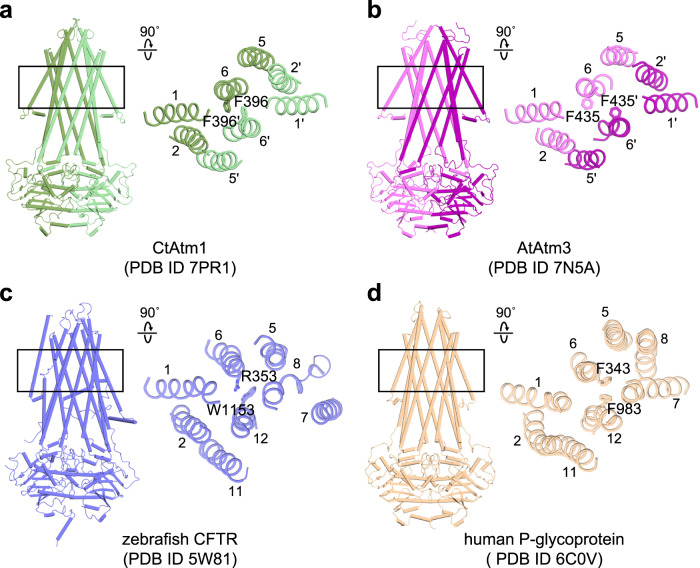
Human ATP-binding cassette transporter subfamily B6 (ABCB6) is a mitochondrial ATP-driven pump that translocates porphyrins from the cytoplasm into mitochondria for heme biosynthesis. Within the transport pathway, a conserved aromatic residue W546 located in each monomer plays a pivotal role in stabilizing the occluded conformation via π-stacking interactions. Herein, we employed cryo-electron microscopy to investigate the structural consequences of a single W546A mutation in ABCB6, both in detergent micelles and nanodiscs. The results demonstrate that the W546A mutation alters the conformational dynamics of detergent-purified ABCB6, leading to entrapment of the transporter in an outward-facing transient state. However, in the nanodisc system, we observed a direct interaction between the transporter and a phospholipid molecule that compensates for the absence of the W546 residue, thereby facilitating the normal conformational transition of the transporter toward the occluded state following ATP hydrolysis. The findings also reveal that adoption of the outward-facing conformation causes charge repulsion between ABCB6 and the bound substrate, and rearrangement of key interacting residues at the substrate-binding site. Consequently, the affinity for the substrate is significantly reduced, facilitating its release from the transporter.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Structural Insights into Porphyrin Recognition by the Human ATP-Binding Cassette Transporter ABCB6.Mol Cells. 2022 Aug 31;45(8):575-587. doi: 10.14348/molcells.2022.0040. Epub 2022 Jul 28. Mol Cells. 2022. PMID: 35950458 Free PMC article.
-
Cryo-electron microscopy structure of human ABCB6 transporter.Protein Sci. 2020 Dec;29(12):2363-2374. doi: 10.1002/pro.3960. Epub 2020 Oct 15. Protein Sci. 2020. PMID: 33007128 Free PMC article.
-
Efficient purification and reconstitution of ATP binding cassette transporter B6 (ABCB6) for functional and structural studies.J Biol Chem. 2013 Aug 2;288(31):22658-69. doi: 10.1074/jbc.M113.485284. Epub 2013 Jun 21. J Biol Chem. 2013. PMID: 23792964 Free PMC article.
-
The role of ABCG2 and ABCB6 in porphyrin metabolism and cell survival.Curr Pharm Biotechnol. 2011 Apr;12(4):647-55. doi: 10.2174/138920111795163995. Curr Pharm Biotechnol. 2011. PMID: 21118089 Review.
-
ABCB6, an ABC Transporter Impacting Drug Response and Disease.AAPS J. 2017 Nov 30;20(1):8. doi: 10.1208/s12248-017-0165-6. AAPS J. 2017. PMID: 29192381 Free PMC article. Review.
Cited by
-
Construction of a Plasmid-Free Escherichia coli Strain with Enhanced Heme Supply to Produce Active Hemoglobins.Metabolites. 2025 Feb 23;15(3):151. doi: 10.3390/metabo15030151. Metabolites. 2025. PMID: 40137115 Free PMC article.
-
ATP-Binding Cassette and Solute Carrier Transporters: Understanding Their Mechanisms and Drug Modulation Through Structural and Modeling Approaches.Pharmaceuticals (Basel). 2024 Nov 27;17(12):1602. doi: 10.3390/ph17121602. Pharmaceuticals (Basel). 2024. PMID: 39770445 Free PMC article. Review.
-
Cryo-EM structure of cadmium-bound human ABCB6.Commun Biol. 2024 May 31;7(1):672. doi: 10.1038/s42003-024-06377-1. Commun Biol. 2024. PMID: 38822018 Free PMC article.
-
A Structural Bioinformatics-Guided Study of Adenosine Triphosphate-Binding Cassette (ABC) Transporters and Their Substrates.Membranes (Basel). 2025 Jan 10;15(1):20. doi: 10.3390/membranes15010020. Membranes (Basel). 2025. PMID: 39852261 Free PMC article.
References
-
- Krishnamurthy PC, et al. Identification of a mammalian mitochondrial porphyrin transporter. Nature. 2006;443:586–589. - PubMed
-
- Tsuchida M, Emi Y, Kida Y, Sakaguchi M. Human ABC transporter isoform B6 (ABCB6) localizes primarily in the Golgi apparatus. Biochem. Biophys. Res. Commun. 2008;369:369–375. - PubMed
-
- Paterson JK, et al. Human ABCB6 localizes to both the outer mitochondrial membrane and the plasma membrane. Biochemistry. 2007;46:9443–9452. - PubMed
-
- Jalil YA, et al. Vesicular localization of the rat ATP-binding cassette half-transporter rAbcb6. Am. J. Physiol. Cell Physiol. 2008;294:C579–C590. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
