

A de novo evolved gene contributes to rice grain shape difference between indica and japonica
- PMID: 37737275
- PMCID: PMC10516980
- DOI: 10.1038/s41467-023-41669-w
A de novo evolved gene contributes to rice grain shape difference between indica and japonica
Abstract
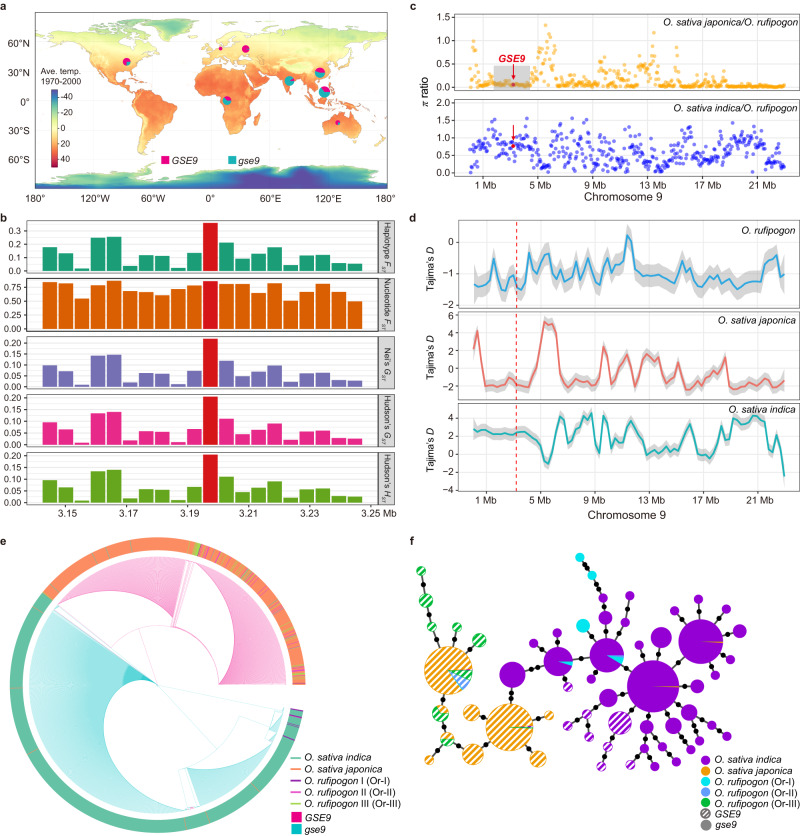
The role of de novo evolved genes from non-coding sequences in regulating morphological differentiation between species/subspecies remains largely unknown. Here, we show that a rice de novo gene GSE9 contributes to grain shape difference between indica/xian and japonica/geng varieties. GSE9 evolves from a previous non-coding region of wild rice Oryza rufipogon through the acquisition of start codon. This gene is inherited by most japonica varieties, while the original sequence (absence of start codon, gse9) is present in majority of indica varieties. Knockout of GSE9 in japonica varieties leads to slender grains, whereas introgression to indica background results in round grains. Population evolutionary analyses reveal that gse9 and GSE9 are derived from wild rice Or-I and Or-III groups, respectively. Our findings uncover that the de novo GSE9 gene contributes to the genetic and morphological divergence between indica and japonica subspecies, and provide a target for precise manipulation of rice grain shape.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures





References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
