

Mass Spectrometric Profiling of HLA-B44 Peptidomes Provides Evidence for Tapasin-Mediated Tryptophan Editing
- PMID: 37737643
- PMCID: PMC10592002
- DOI: 10.4049/jimmunol.2300232
Mass Spectrometric Profiling of HLA-B44 Peptidomes Provides Evidence for Tapasin-Mediated Tryptophan Editing
Abstract
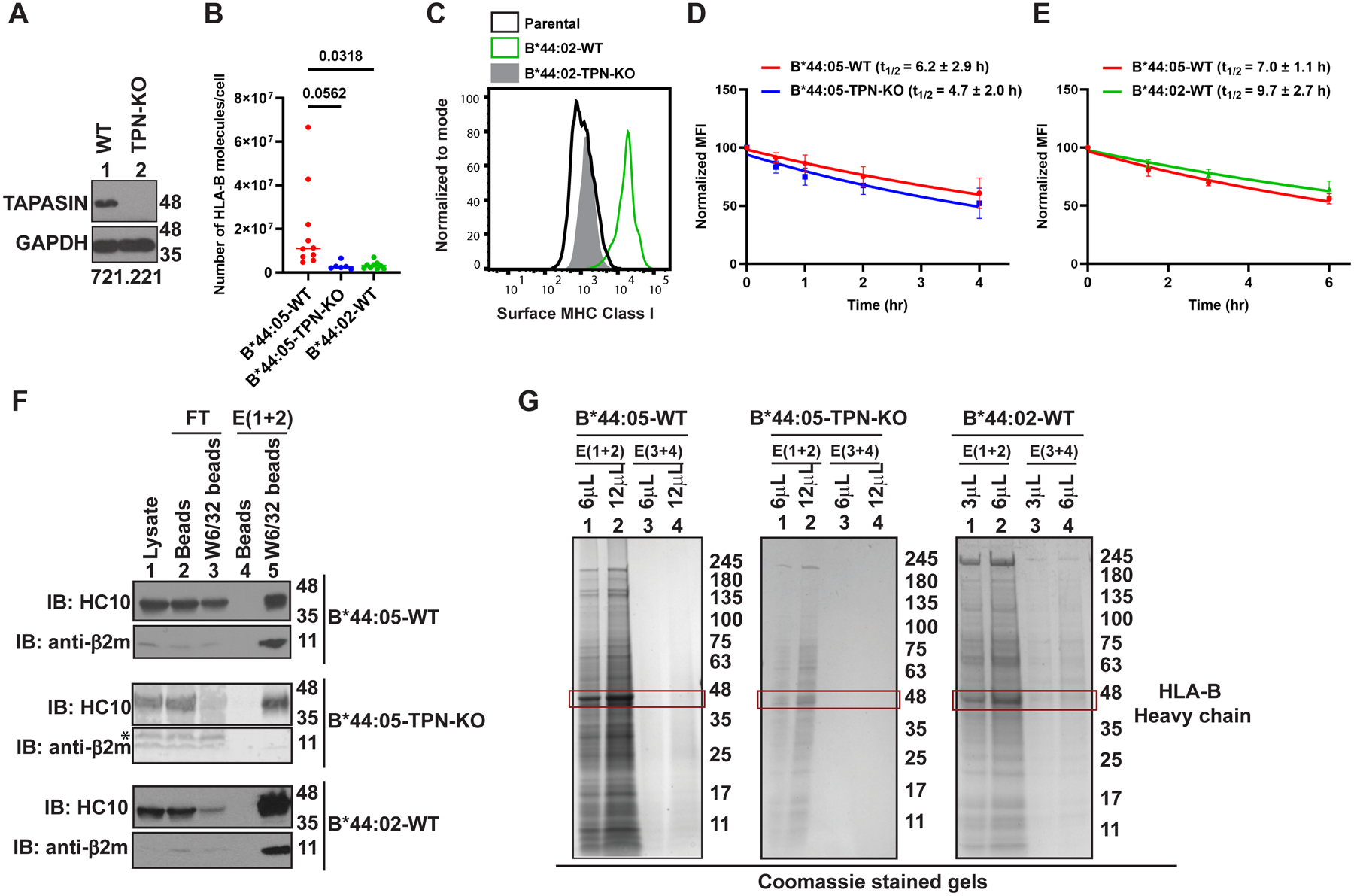
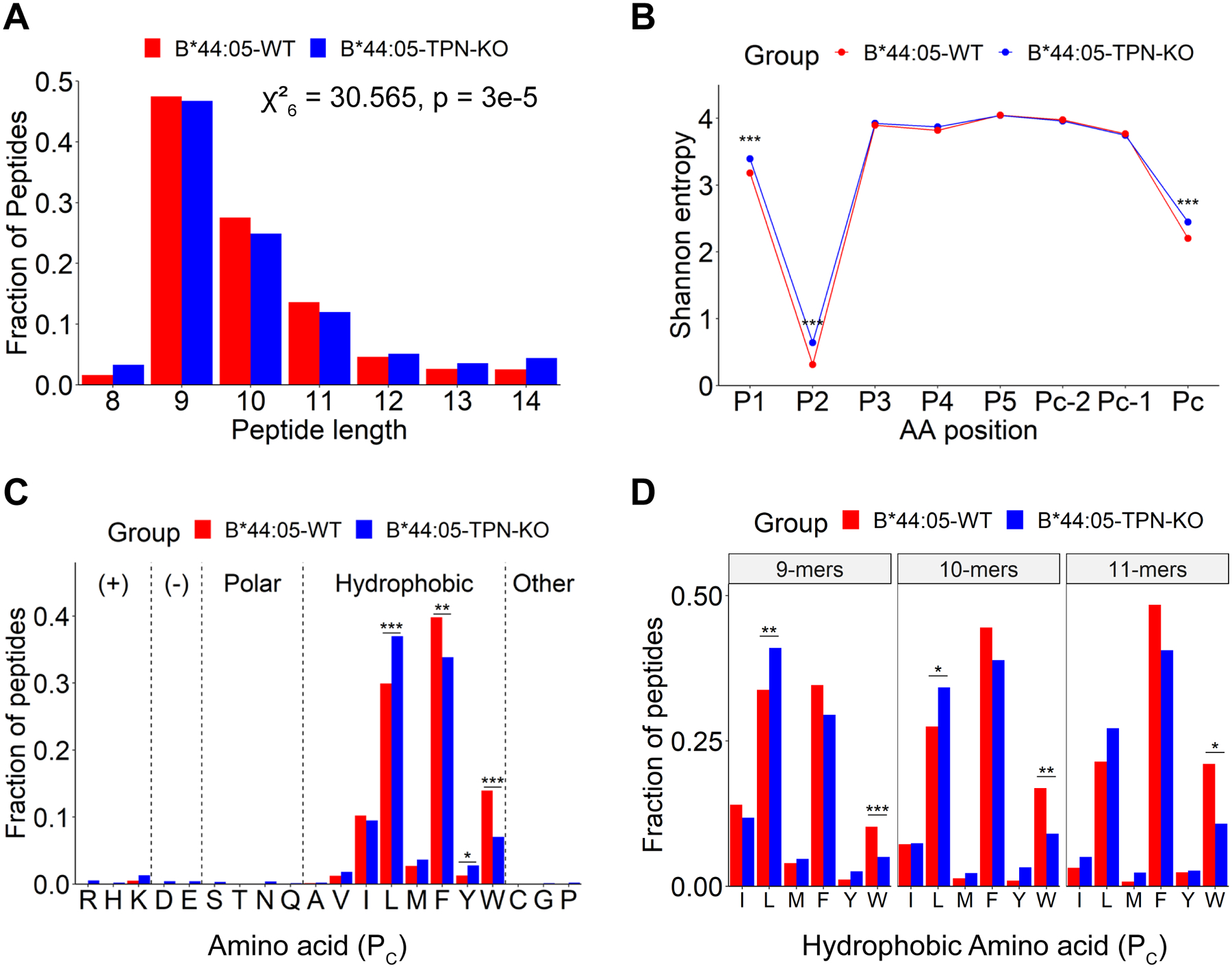
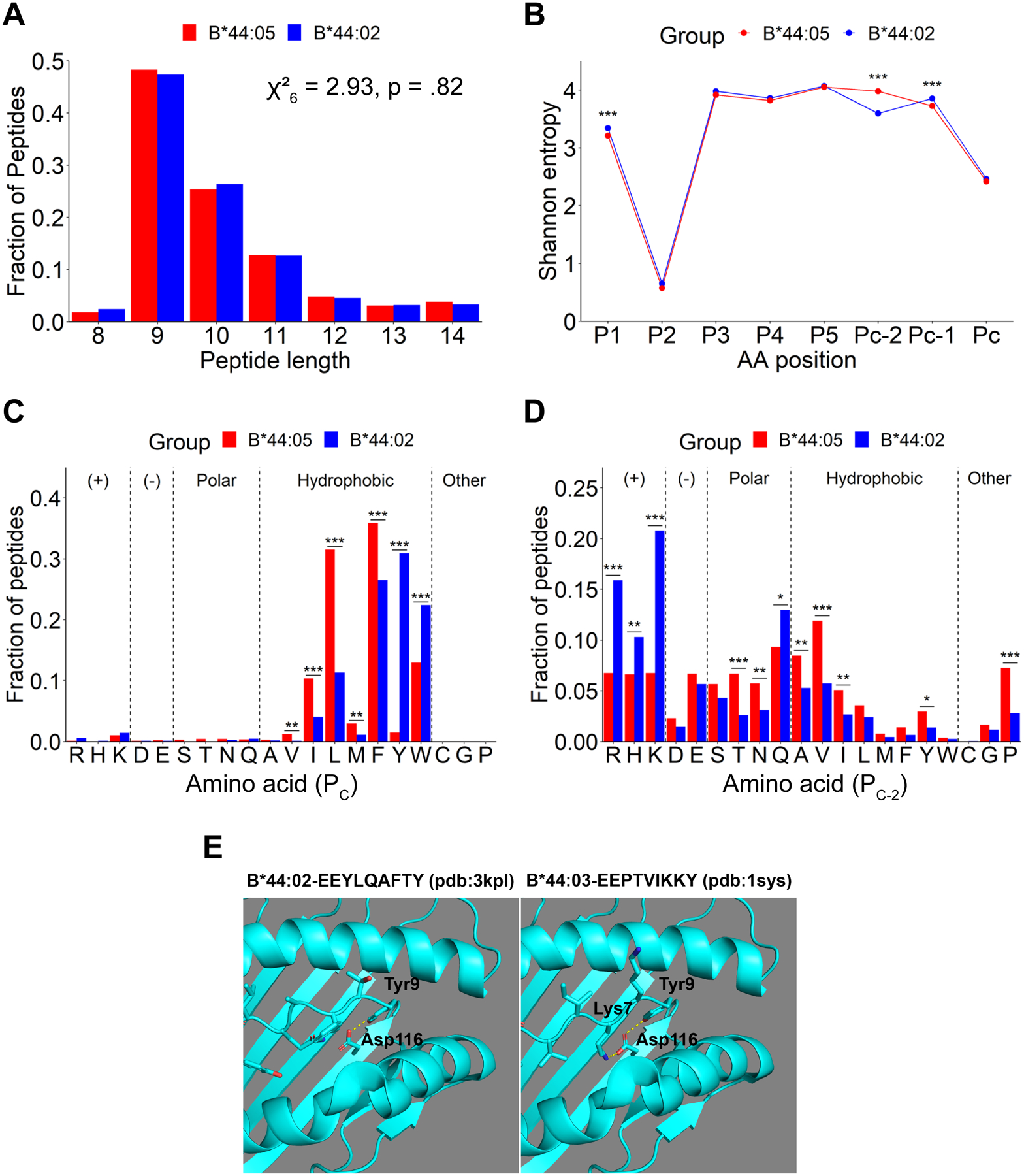
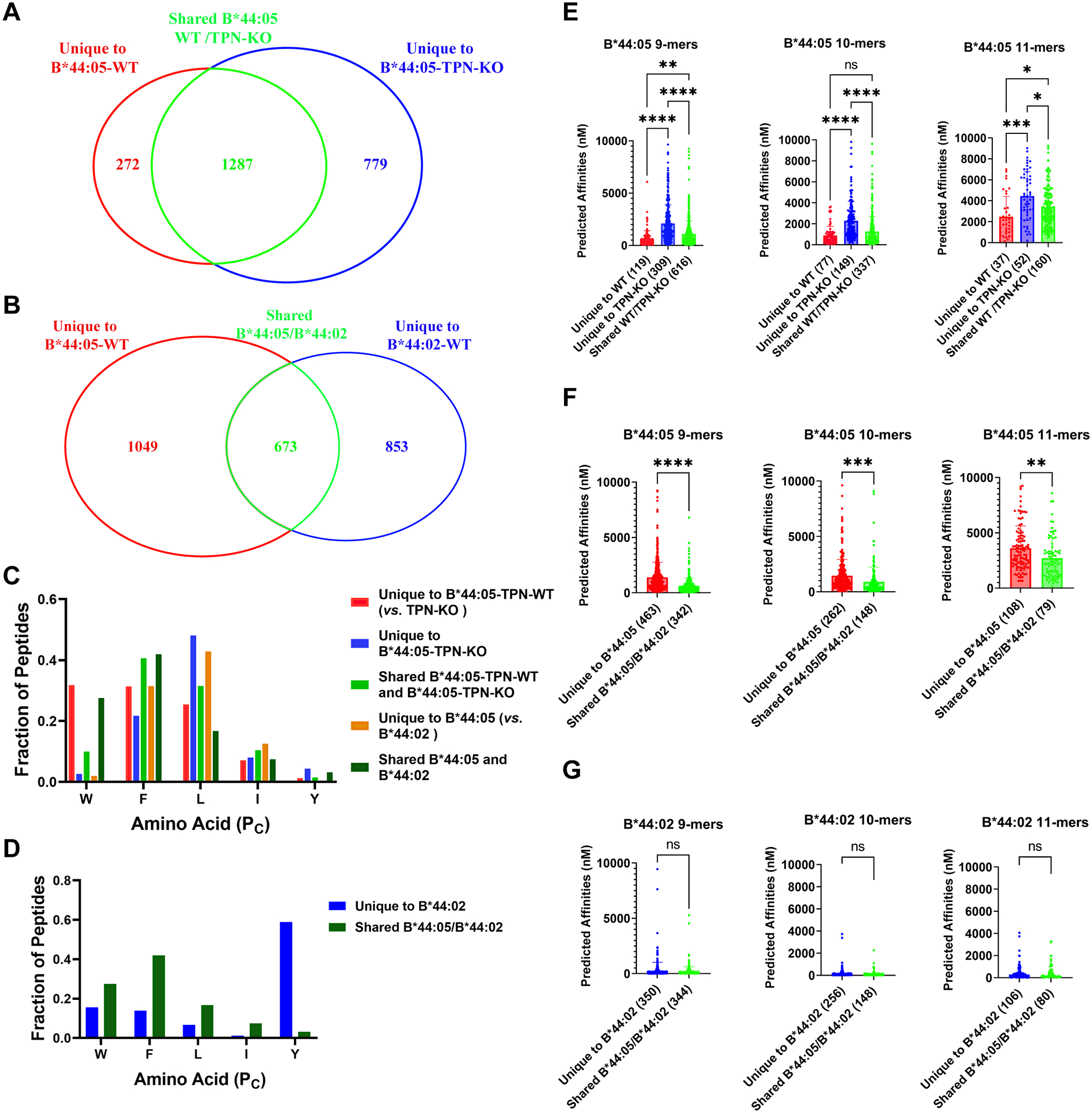
The extreme polymorphisms of HLA class I proteins result in structural variations in their peptide binding sites to achieve diversity in Ag presentation. External factors could independently constrict or alter HLA class I peptide repertoires. Such effects of the assembly factor tapasin were assessed for HLA-B*44:05 (Y116) and a close variant, HLA-B*44:02 (D116), which have low and high tapasin dependence, respectively, for their cell surface expression. Analyses of the HLA-B*44:05 peptidomes in the presence and absence of tapasin reveal that peptides with C-terminal tryptophans and higher predicted affinities are preferentially selected by tapasin, coincident with reduced frequencies of peptides with other C-terminal amino acids, including leucine. Comparisons of the HLA-B*44:05 and HLA-B*44:02 peptidomes indicate the expected structure-based alterations near the peptide C termini, but also C-terminal amino acid frequency and predicted affinity changes among the unique and shared peptide groups for B*44:02 and B*44:05. Overall, these findings indicate that the presence of tapasin and the tapasin dependence of assembly alter HLA class I peptide-binding preferences at the peptide C terminus. The particular C-terminal amino acid preferences that are altered by tapasin are expected to be determined by the intrinsic peptide-binding specificities of HLA class I allotypes. Additionally, the findings suggest that tapasin deficiency and reduced tapasin dependence expand the permissive affinities of HLA class I-bound peptides, consistent with prior findings that HLA class I allotypes with low tapasin dependence have increased breadth of CD8+ T cell epitope presentation and are more protective in HIV infections.
Copyright © 2023 by The American Association of Immunologists, Inc.
Conflict of interest statement
Conflicts of Interest
All the authors have no conflicts of interest to declare.
Figures
Update of
-
Mass spectrometric profiling of HLA-B44 peptidomes provides evidence for tapasin-mediated tryptophan editing.bioRxiv [Preprint]. 2023 Feb 27:2023.02.26.530125. doi: 10.1101/2023.02.26.530125. bioRxiv. 2023. Update in: J Immunol. 2023 Nov 1;211(9):1298-1307. doi: 10.4049/jimmunol.2300232. PMID: 36909546 Free PMC article. Updated. Preprint.
References
-
- Jiang J, Natarajan K, and Margulies DH. 2019. MHC Molecules, T cell Receptors, Natural Killer Cell Receptors, and Viral Immunoevasins-Key Elements of Adaptive and Innate Immunity. Adv Exp Med Biol 1172: 21–62. - PubMed
-
- Blees A, Januliene D, Hofmann T, Koller N, Schmidt C, Trowitzsch S, Moeller A, and Tampe R. 2017. Structure of the human MHC-I peptide-loading complex. Nature 551: 525–528. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
