

Evolutionarily conserved protein motifs drive interactions between the plant nucleoskeleton and nuclear pores
- PMID: 37738557
- PMCID: PMC10689174
- DOI: 10.1093/plcell/koad236
Evolutionarily conserved protein motifs drive interactions between the plant nucleoskeleton and nuclear pores
Abstract
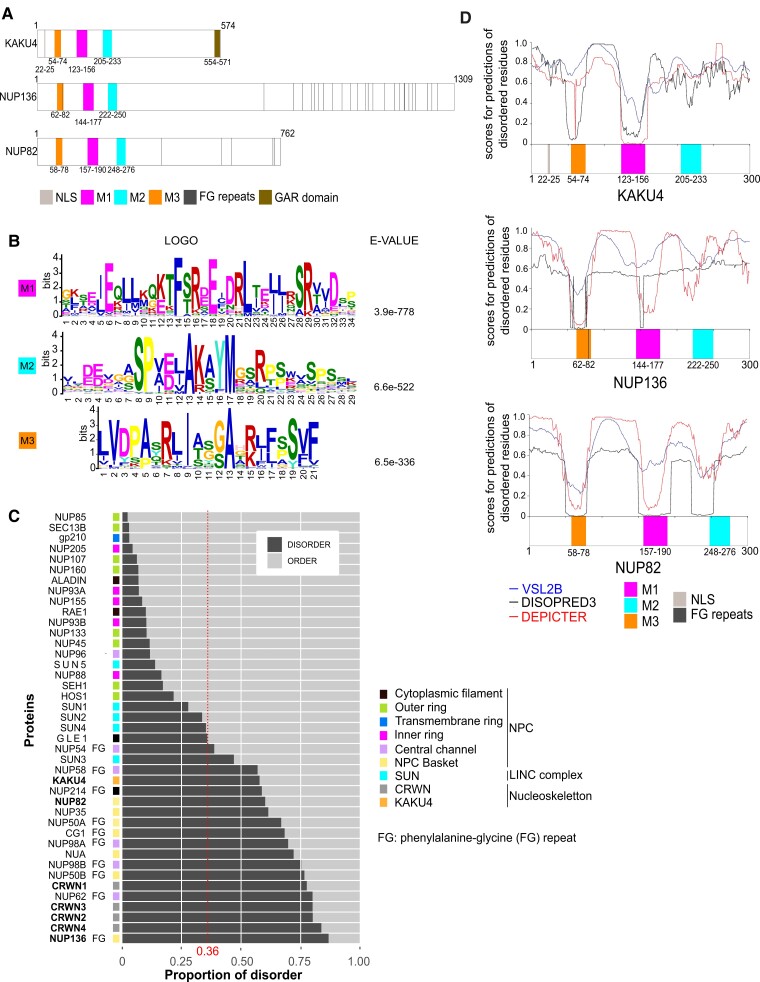
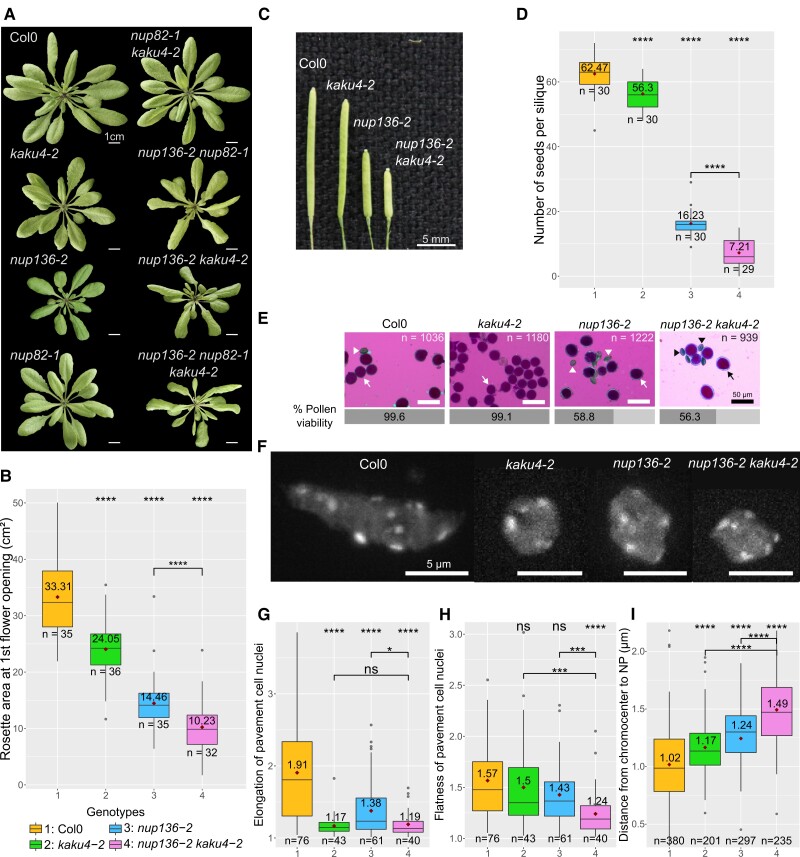
The nucleoskeleton forms a filamentous meshwork under the nuclear envelope and contributes to the regulation of nuclear shape and gene expression. To understand how the Arabidopsis (Arabidopsis thaliana) nucleoskeleton physically connects to the nuclear periphery in plants, we investigated the Arabidopsis nucleoskeleton protein KAKU4 and sought for functional regions responsible for its localization at the nuclear periphery. We identified 3 conserved peptide motifs within the N-terminal region of KAKU4, which are required for intermolecular interactions of KAKU4 with itself, interaction with the nucleoskeleton protein CROWDED NUCLEI (CRWN), localization at the nuclear periphery, and nuclear elongation in differentiated tissues. Unexpectedly, we find these motifs to be present also in NUP82 and NUP136, 2 plant-specific nucleoporins from the nuclear pore basket. We further show that NUP82, NUP136, and KAKU4 have a common evolutionary history predating nonvascular land plants with KAKU4 mainly localizing outside the nuclear pore suggesting its divergence from an ancient nucleoporin into a new nucleoskeleton component. Finally, we demonstrate that both NUP82 and NUP136, through their shared N-terminal motifs, interact with CRWN and KAKU4 proteins revealing the existence of a physical continuum between the nuclear pore and the nucleoskeleton in plants.
© American Society of Plant Biologists 2023. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Conflict of interest statement
Conflict of interest statement. None declared.
Figures





Similar articles
-
GBPL3 localizes to the nuclear pore complex and functionally connects the nuclear basket with the nucleoskeleton in plants.PLoS Biol. 2022 Oct 21;20(10):e3001831. doi: 10.1371/journal.pbio.3001831. eCollection 2022 Oct. PLoS Biol. 2022. PMID: 36269771 Free PMC article.
-
Nup82 functions redundantly with Nup136 in a salicylic acid-dependent defense response of Arabidopsis thaliana.Nucleus. 2017 May 4;8(3):301-311. doi: 10.1080/19491034.2017.1279774. Epub 2017 Jan 10. Nucleus. 2017. PMID: 28071978 Free PMC article.
-
Involvement of the nuclear pore complex in morphology of the plant nucleus.Nucleus. 2011 May-Jun;2(3):168-72. doi: 10.4161/nucl.2.3.16175. Nucleus. 2011. PMID: 21818409 Free PMC article.
-
The plant LINC complex at the nuclear envelope.Chromosome Res. 2014 Jun;22(2):241-52. doi: 10.1007/s10577-014-9419-7. Chromosome Res. 2014. PMID: 24801343 Review.
-
Assessing the function of the plant nuclear pore complex and the search for specificity.J Exp Bot. 2013 Feb;64(4):833-45. doi: 10.1093/jxb/ers289. Epub 2012 Oct 17. J Exp Bot. 2013. PMID: 23077202 Review.
Cited by
-
CRWN nuclear lamina components maintain the H3K27me3 landscape and promote successful reproduction in Arabidopsis.New Phytol. 2024 Jul;243(1):213-228. doi: 10.1111/nph.19791. Epub 2024 May 7. New Phytol. 2024. PMID: 38715414 Free PMC article.
-
GBPL3 localizes to the nuclear pore complex and functionally connects the nuclear basket with the nucleoskeleton in plants.PLoS Biol. 2022 Oct 21;20(10):e3001831. doi: 10.1371/journal.pbio.3001831. eCollection 2022 Oct. PLoS Biol. 2022. PMID: 36269771 Free PMC article.
-
The N-terminal coiled-coil domain of Arabidopsis CROWDED NUCLEI 1 is required for nuclear morphology maintenance.Planta. 2024 Jul 27;260(3):62. doi: 10.1007/s00425-024-04489-w. Planta. 2024. PMID: 39066892
-
Nuclear lamina phase separation orchestrates stress-induced transcriptional responses in plants.Dev Cell. 2025 Aug 1:S1534-5807(25)00446-0. doi: 10.1016/j.devcel.2025.07.008. Online ahead of print. Dev Cell. 2025. PMID: 40780209 Free PMC article.
-
Plant nuclear envelope as a hub connecting genome organization with regulation of gene expression.Nucleus. 2023 Dec;14(1):2178201. doi: 10.1080/19491034.2023.2178201. Nucleus. 2023. PMID: 36794966 Free PMC article.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
