

Impact of SARS-CoV-2 ORF6 and its variant polymorphisms on host responses and viral pathogenesis
- PMID: 37738983
- PMCID: PMC10750313
- DOI: 10.1016/j.chom.2023.08.003
Impact of SARS-CoV-2 ORF6 and its variant polymorphisms on host responses and viral pathogenesis
Abstract
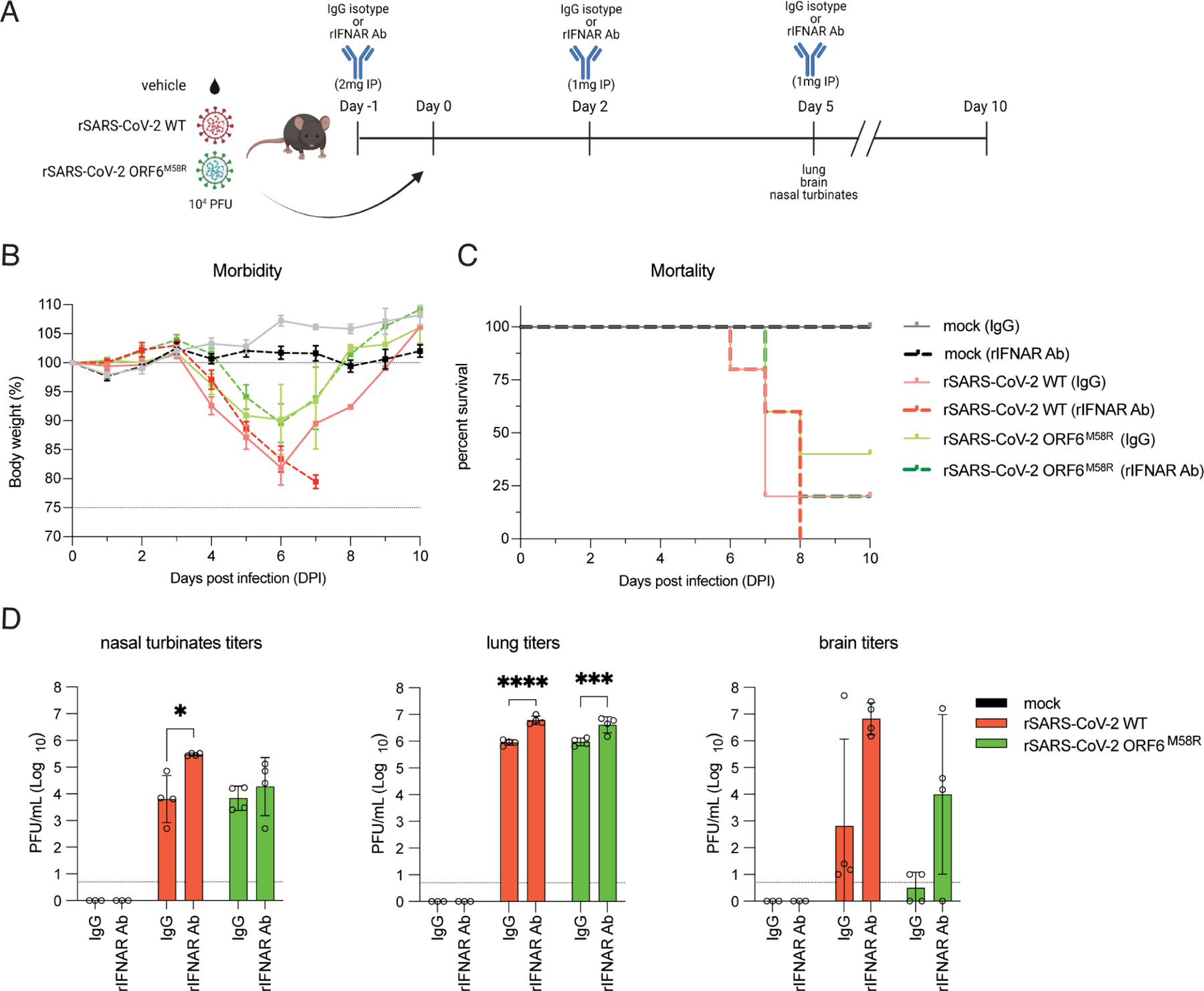
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) encodes several proteins that inhibit host interferon responses. Among these, ORF6 antagonizes interferon signaling by disrupting nucleocytoplasmic trafficking through interactions with the nuclear pore complex components Nup98-Rae1. However, the roles and contributions of ORF6 during physiological infection remain unexplored. We assessed the role of ORF6 during infection using recombinant viruses carrying a deletion or loss-of-function (LoF) mutation in ORF6. ORF6 plays key roles in interferon antagonism and viral pathogenesis by interfering with nuclear import and specifically the translocation of IRF and STAT transcription factors. Additionally, ORF6 inhibits cellular mRNA export, resulting in the remodeling of the host cell proteome, and regulates viral protein expression. Interestingly, the ORF6:D61L mutation that emerged in the Omicron BA.2 and BA.4 variants exhibits reduced interactions with Nup98-Rae1 and consequently impairs immune evasion. Our findings highlight the role of ORF6 in antagonizing innate immunity and emphasize the importance of studying the immune evasion strategies of SARS-CoV-2.
Keywords: ORF6; Omicron variant; SARS-CoV-2; SARS-CoV-2 pathogenesis; interferon; mRNA export; nuclear import; nucleocytoplasmic trafficking; virus-host interaction.
Copyright © 2023 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The A.G.-S. laboratory has received research support from Pfizer, Senhwa Biosciences, Kenall Manufacturing, Blade Therapuetics, Avimex, Johnson & Johnson, Dynavax, 7Hills Pharma, Pharmamar, ImmunityBio, Accurius, Nanocomposix, Hexamer, N-fold LLC, Model Medicines, Atea Pharma, Applied Biological Laboratories and Merck, outside of the reported work. A.G.-S. has consulting agreements for the following companies involving cash and/or stock: Castlevax, Amovir, Vivaldi Biosciences, Contrafect, 7Hills Pharma, Avimex, Vaxalto, Pagoda, Accurius, Esperovax, Farmak, Applied Biological Laboratories, Pharmamar, Paratus, CureLab Oncology, CureLab Veterinary, Synairgen and Pfizer, outside of the reported work. A.G.-S. has been an invited speaker in meeting events organized by Seqirus, Janssen, Abbott, and Astrazeneca. A.G.-S. is inventor on patents and patent applications on the use of antivirals and vaccines for the treatment and prevention of virus infections and cancer, owned by ISMMS, New York. C.Y. and L. M.-S. are co-inventors on a patent application directed to reverse genetics approaches to generate recombinant SARS-CoV-2. The Krogan Laboratory has received research support from Vir Biotechnology, F. Hoffmann-La Roche, and Rezo Therapeutics. N.J.K. has previously held financially compensated consulting agreements with the Icahn School of Medicine at Mount Sinai, New York and Twist Bioscience Corp. He currently has financially compensated consulting agreements with Maze Therapeutics, Interline Therapeutics, Rezo Therapeutics, and GEn1E Lifesciences, Inc.. He is on the Board of Directors of Rezo Therapeutics and is a shareholder in Tenaya Therapeutics, Maze Therapeutics, Rezo Therapeutics, and Interline Therapeutics. A.M. is the creator of Omics Bioinformatics and owns all the stocks of this company. ISMMS has filed patent applications relating to SARS-CoV-2 serological assays which list V.S. as co-inventor.
Figures






Update of
-
Impact of SARS-CoV-2 ORF6 and its variant polymorphisms on host responses and viral pathogenesis.bioRxiv [Preprint]. 2022 Nov 30:2022.10.18.512708. doi: 10.1101/2022.10.18.512708. bioRxiv. 2022. Update in: Cell Host Microbe. 2023 Oct 11;31(10):1668-1684.e12. doi: 10.1016/j.chom.2023.08.003. PMID: 36299428 Free PMC article. Updated. Preprint.
References
-
- Boudewijns R, Thibaut HJ, Kaptein SJF, Li R, Vergote V, Seldeslachts L, Van Weyenbergh J, De Keyzer C, Bervoets L, Sharma S, et al. (2020). STAT2 signaling restricts viral dissemination but drives severe pneumonia in SARS-CoV-2 infected hamsters. Nat Commun 11, 5838. 10.1038/s41467-020-19684-y. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
- K99 AI163868/AI/NIAID NIH HHS/United States
- U19 AI142733/AI/NIAID NIH HHS/United States
- UL1 TR004419/TR/NCATS NIH HHS/United States
- U19 AI171110/AI/NIAID NIH HHS/United States
- R01 AI151029/AI/NIAID NIH HHS/United States
- R01 AI161175/AI/NIAID NIH HHS/United States
- U54 CA260560/CA/NCI NIH HHS/United States
- T32 AI007647/AI/NIAID NIH HHS/United States
- R21 AI173816/AI/NIAID NIH HHS/United States
- S10 OD026983/OD/NIH HHS/United States
- U19 AI135990/AI/NIAID NIH HHS/United States
- S10 OD030269/OD/NIH HHS/United States
- R01 AI161363/AI/NIAID NIH HHS/United States
- U19 AI135972/AI/NIAID NIH HHS/United States
- 75N93021C00014/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
