

Sequence tolerance of immunoglobulin variable domain framework regions to noncanonical intradomain disulfide linkages
- PMID: 37742917
- PMCID: PMC10641266
- DOI: 10.1016/j.jbc.2023.105278
Sequence tolerance of immunoglobulin variable domain framework regions to noncanonical intradomain disulfide linkages
Abstract
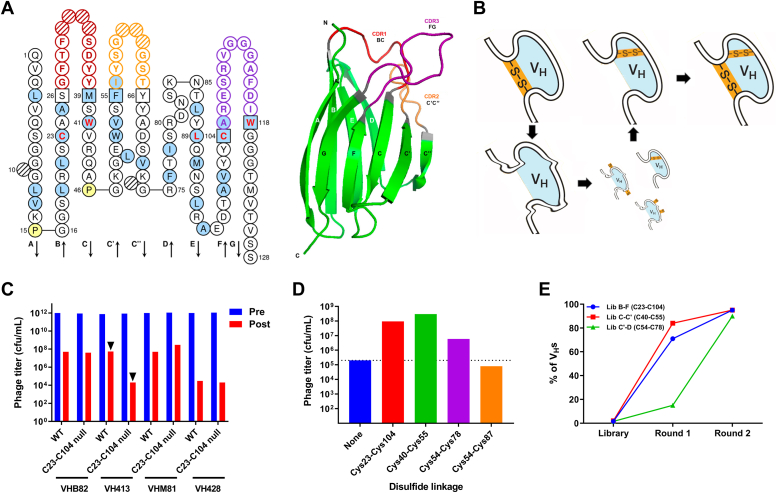
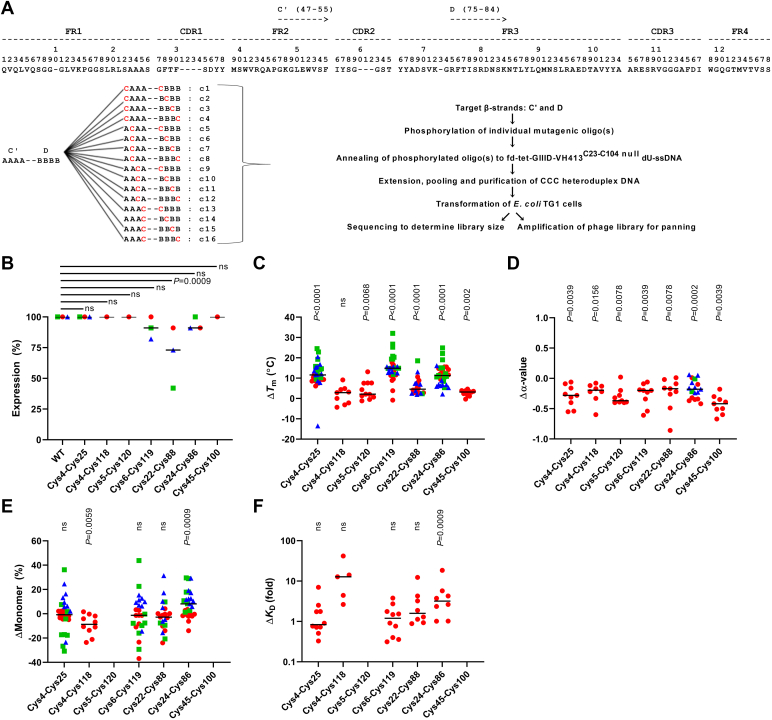
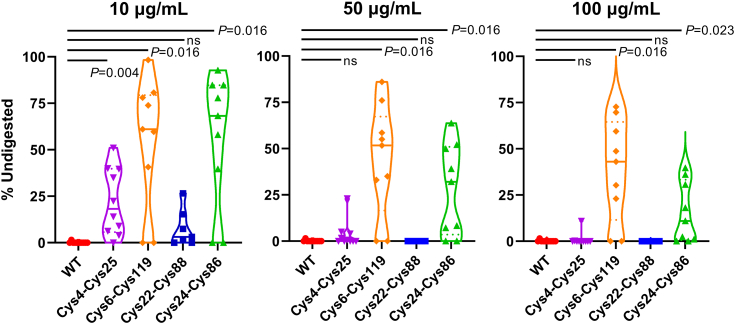
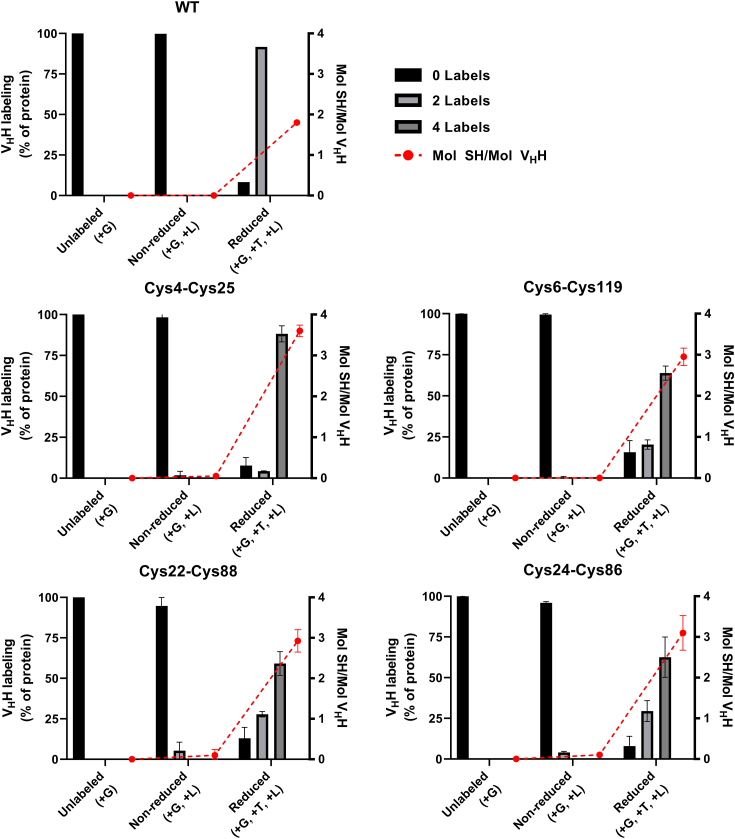
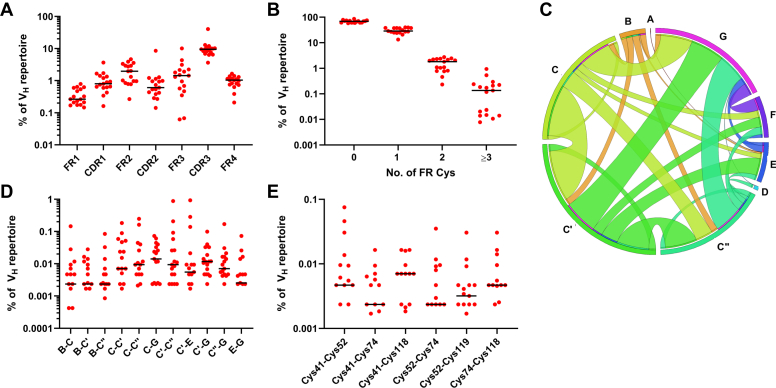
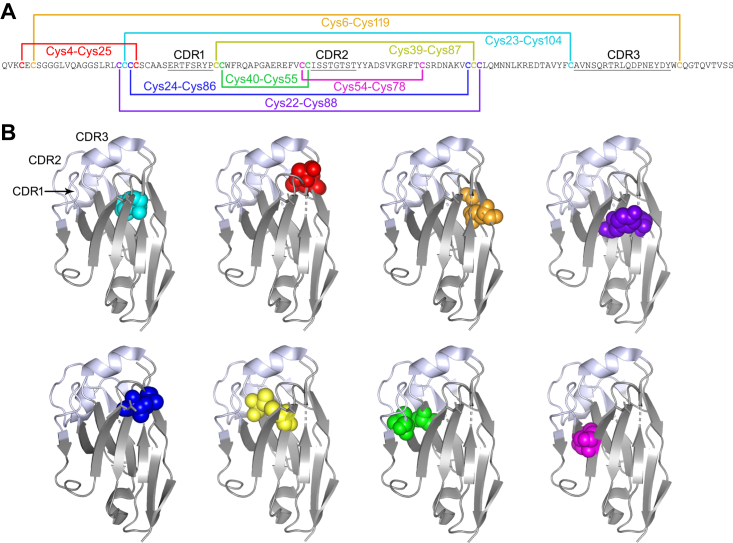
Most immunoglobulin (Ig) domains bear only a single highly conserved canonical intradomain, inter-β-sheet disulfide linkage formed between Cys23-Cys104, and incorporation of rare noncanonical disulfide linkages at other locations can enhance Ig domain stability. Here, we exhaustively surveyed the sequence tolerance of Ig variable (V) domain framework regions (FRs) to noncanonical disulfide linkages. Starting from a destabilized VH domain lacking a Cys23-Cys104 disulfide linkage, we generated and screened phage-displayed libraries of engineered VHs, bearing all possible pairwise combinations of Cys residues in neighboring β-strands of the Ig fold FRs. This approach identified seven novel Cys pairs in VH FRs (Cys4-Cys25, Cys4-Cys118, Cys5-Cys120, Cys6-Cys119, Cys22-Cys88, Cys24-Cys86, and Cys45-Cys100; the international ImMunoGeneTics information system numbering), whose presence rescued domain folding and stability. Introduction of a subset of these noncanonical disulfide linkages (three intra-β-sheet: Cys4-Cys25, Cys22-Cys88, and Cys24-Cys86, and one inter-β-sheet: Cys6-Cys119) into a diverse panel of VH, VL, and VHH domains enhanced their thermostability and protease resistance without significantly impacting expression, solubility, or binding to cognate antigens. None of the noncanonical disulfide linkages identified were present in the natural human VH repertoire. These data reveal an unexpected permissiveness of Ig V domains to noncanonical disulfide linkages at diverse locations in FRs, absent in the human repertoire, whose presence is compatible with antigen recognition and improves domain stability. Our work represents the most complete assessment to date of the role of engineered noncanonical disulfide bonding within FRs in Ig V domain structure and function.
Keywords: disulfide; immunoglobulin variable domain; phage display; protein engineering; protein stability; single-domain antibody (sdAb, nanobody).
Crown Copyright © 2023. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Dombkowski A.A., Sultana K.Z., Craig D.B. Protein disulfide engineering. FEBS Lett. 2013;588:206–212. - PubMed
-
- Willuda J., Honegger A., Waibel R., Schubiger P.A., Stahel R., Zangemeister-Wittke U., et al. Thermal stability is essential for tumor targeting of antibody fragments: engineering of a humanized anti-epithelial glycoprotein-2 (epithelial cell adhesion molecule) single-chain Fv fragment. Cancer Res. 1999;59:5758–5767. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
