

Convergent TP53 loss and evolvability in cancer
- PMID: 37743495
- PMCID: PMC10518978
- DOI: 10.1186/s12862-023-02146-6
Convergent TP53 loss and evolvability in cancer
Abstract
Cancer cell populations evolve by a stepwise process involving natural selection of the fittest variants within a tissue ecosystem context and as modified by therapy. Genomic scrutiny of patient samples reveals an extraordinary diversity of mutational profiles both between patients with similar cancers and within the cancer cell population of individual patients. Does this signify highly divergent evolutionary trajectories or are there repetitive and predictable patterns?Major evolutionary innovations or adaptations in different species are frequently repeated, or convergent, reflecting both common selective pressures and constraints on optimal solutions. We argue this is true of evolving cancer cells, especially with respect to the TP53 gene. Functional loss variants in TP53 are the most common genetic change in cancer. We discuss the likely microenvironmental selective pressures involved and the profound impact this has on cell fitness, evolvability and probability of subsequent drug resistance.
Keywords: TP53; cancer; convergence; drug resistance; hypoxia; stem cells.
© 2023. BioMed Central Ltd., part of Springer Nature.
Conflict of interest statement
The authors declare no competing interests.
Figures
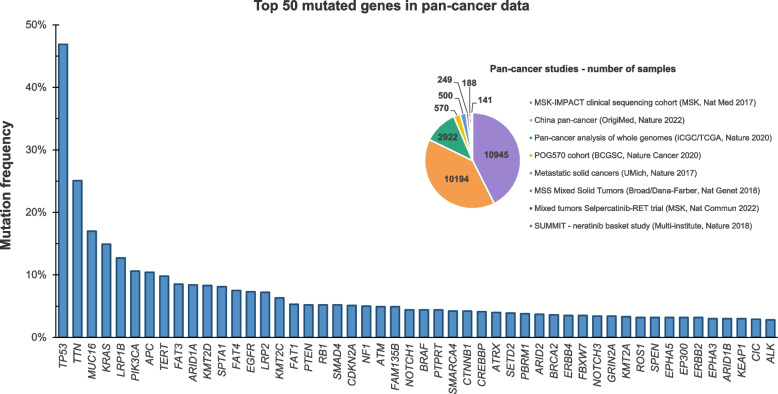
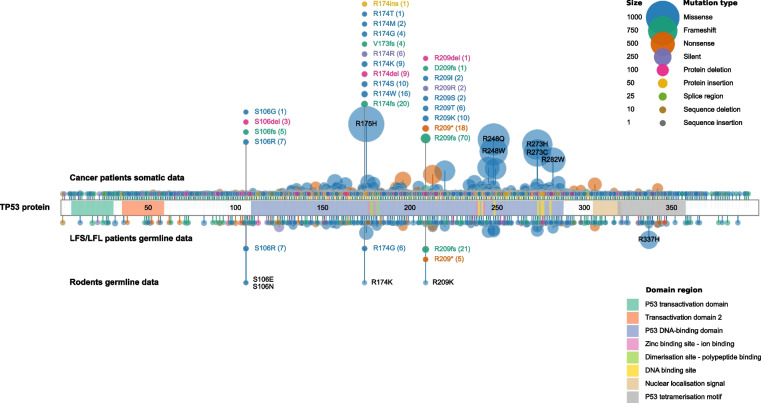
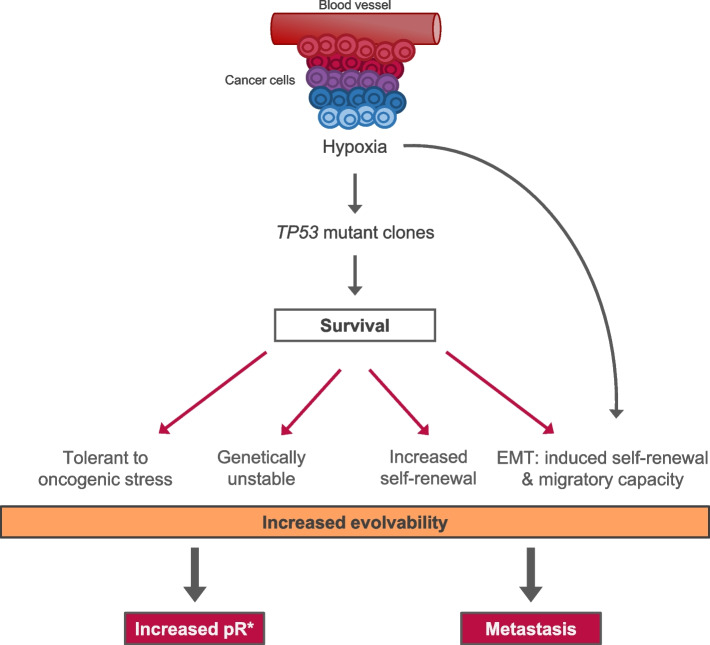
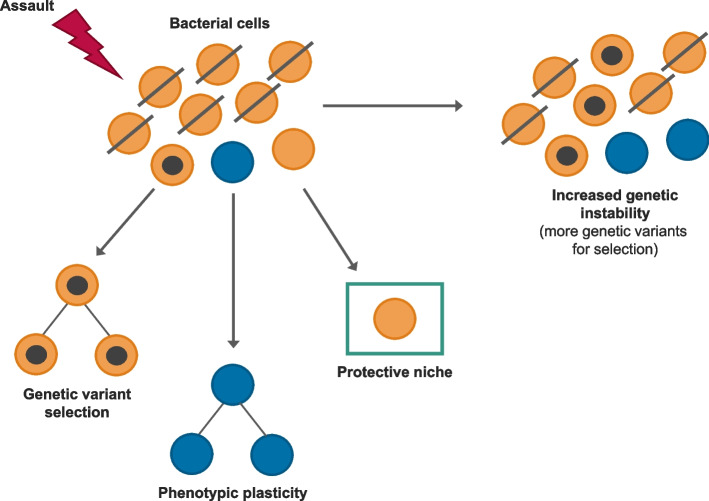
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous