

Biosynthesis of Fusapyrone Depends on the H3K9 Methyltransferase, FmKmt1, in Fusarium mangiferae
- PMID: 37744112
- PMCID: PMC10512364
- DOI: 10.3389/ffunb.2021.671796
Biosynthesis of Fusapyrone Depends on the H3K9 Methyltransferase, FmKmt1, in Fusarium mangiferae
Abstract
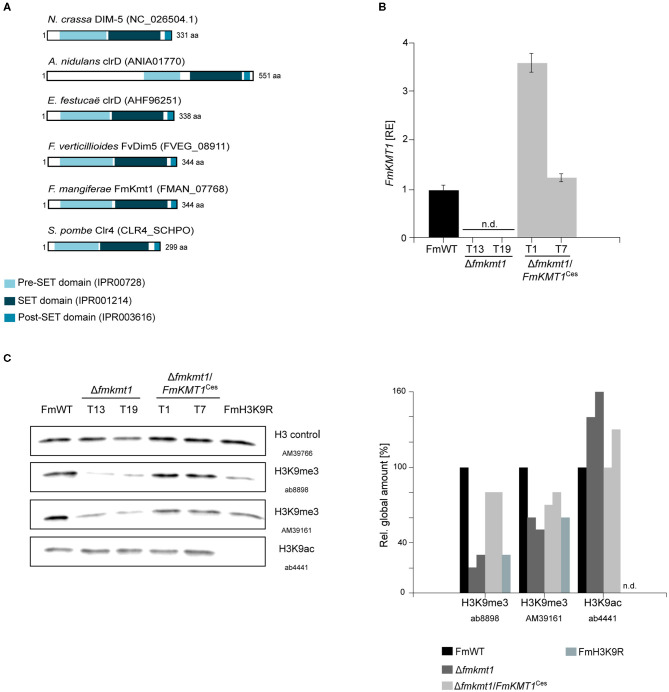
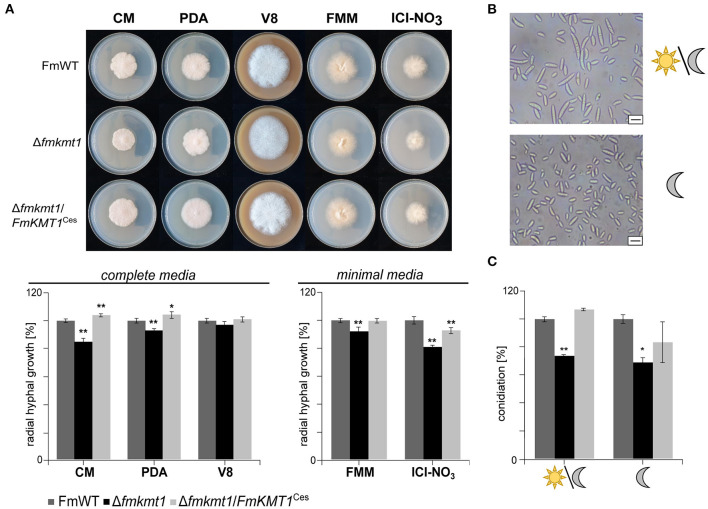
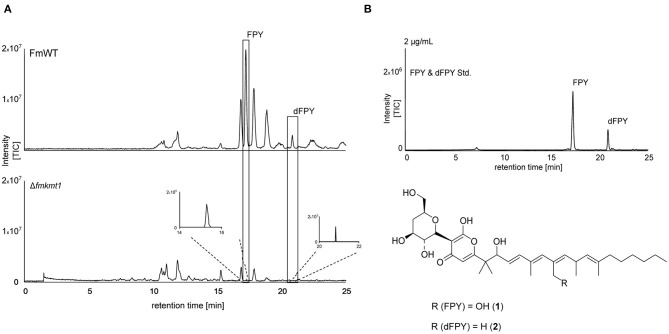
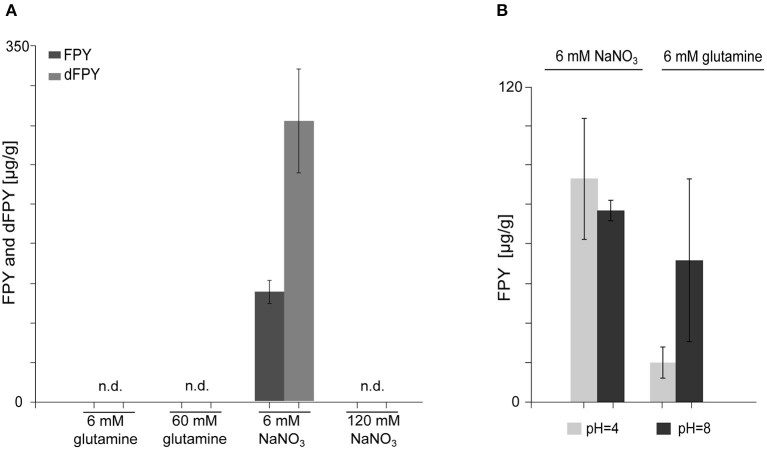
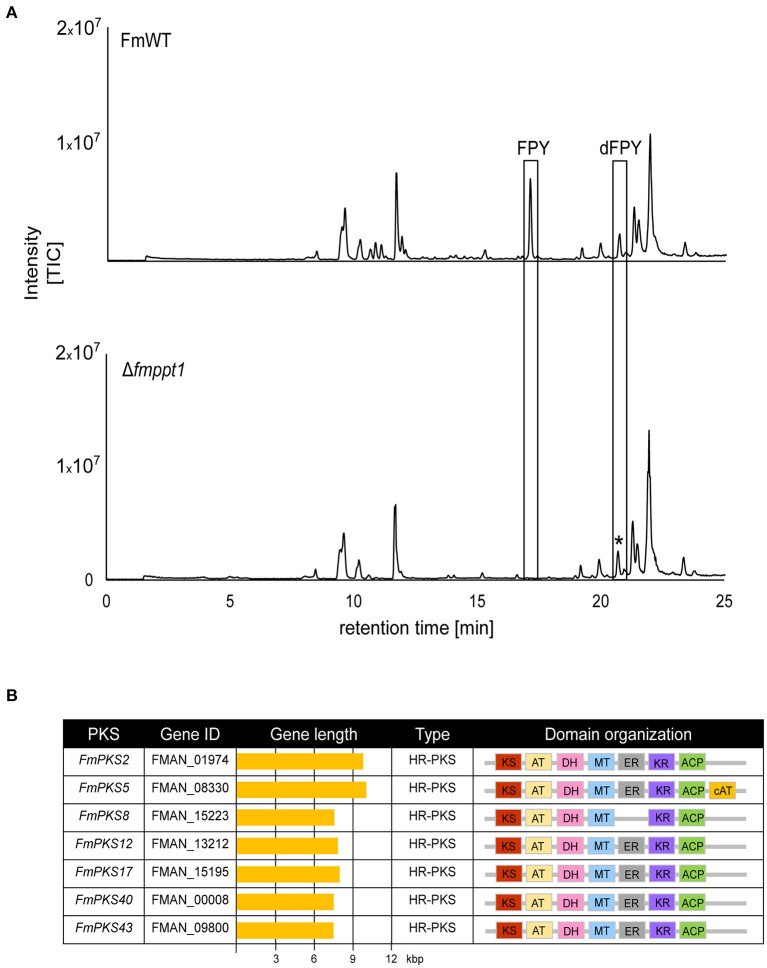
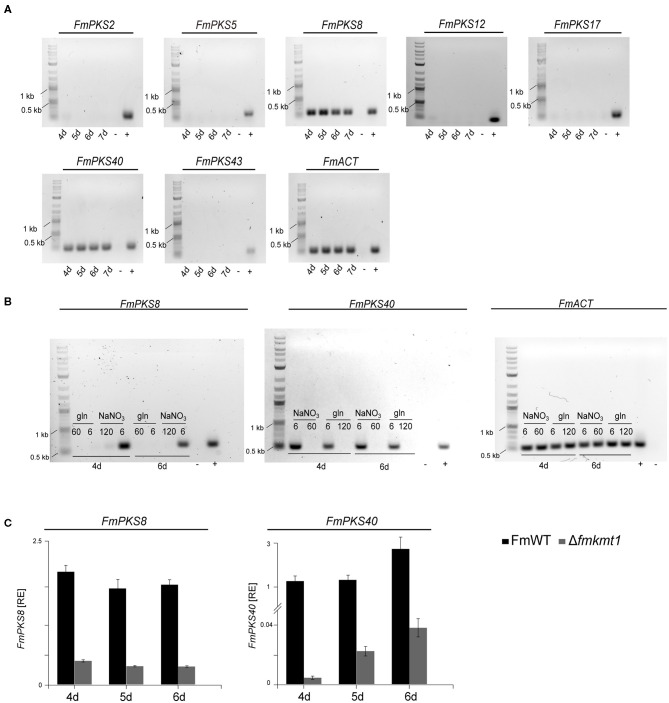
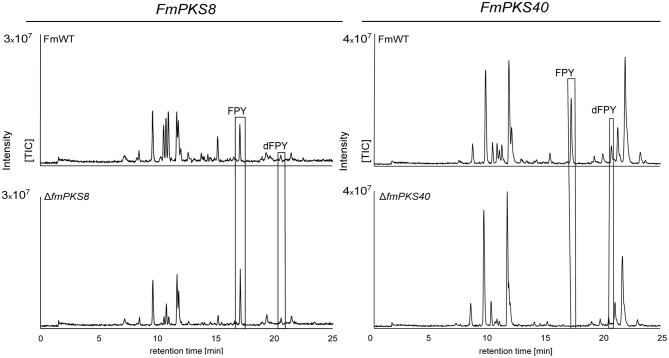
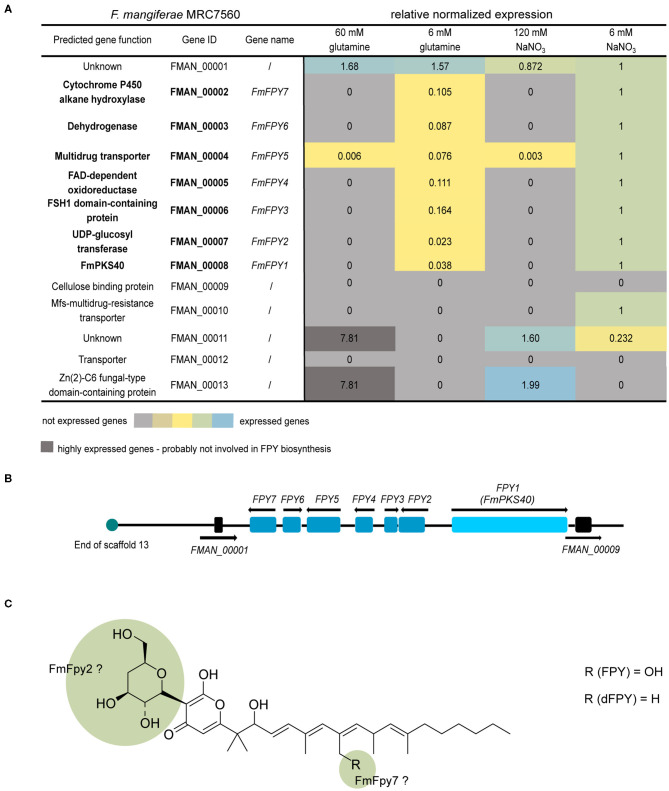
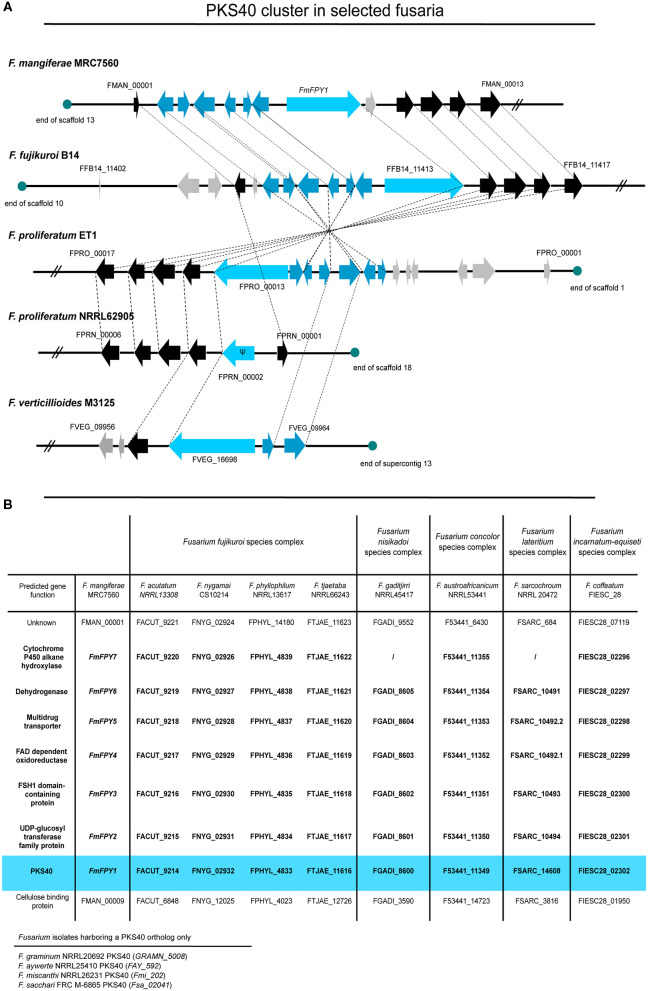
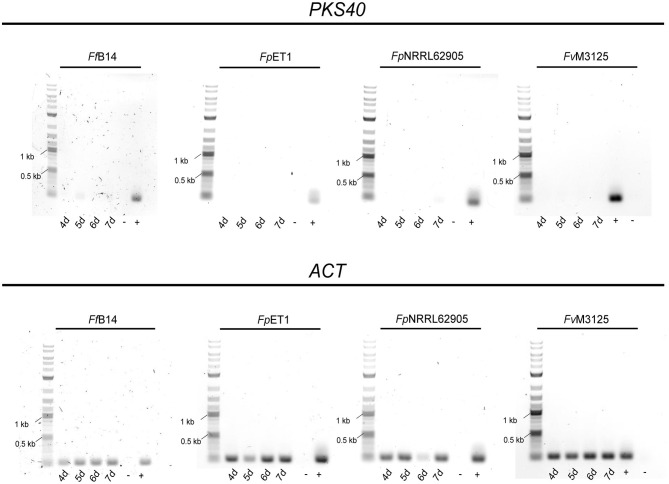
The phytopathogenic fungus Fusarium mangiferae belongs to the Fusarium fujikuroi species complex (FFSC). Members of this group cause a wide spectrum of devastating diseases on diverse agricultural crops. F. mangiferae is the causal agent of the mango malformation disease (MMD) and as such detrimental for agriculture in the southern hemisphere. During plant infection, the fungus produces a plethora of bioactive secondary metabolites (SMs), which most often lead to severe adverse defects on plants health. Changes in chromatin structure achieved by posttranslational modifications (PTM) of histones play a key role in regulation of fungal SM biosynthesis. Posttranslational tri-methylation of histone 3 lysine 9 (H3K9me3) is considered a hallmark of heterochromatin and established by the SET-domain protein Kmt1. Here, we show that FmKmt1 is involved in H3K9me3 in F. mangiferae. Loss of FmKmt1 only slightly though significantly affected fungal hyphal growth and stress response and is required for wild type-like conidiation. While FmKmt1 is largely dispensable for the biosynthesis of most known SMs, removal of FmKMT1 resulted in an almost complete loss of fusapyrone and deoxyfusapyrone, γ-pyrones previously only known from Fusarium semitectum. Here, we identified the polyketide synthase (PKS) FmPKS40 to be involved in fusapyrone biosynthesis, delineate putative cluster borders by co-expression studies and provide insights into its regulation.
Keywords: Fusarium fujikuroi species complex; Fusarium mangiferae; H3K9me3; deoxyfusapyrone; fusapyrone; heterochromatin; histone PTMs; secondary metabolism.
Copyright © 2021 Atanasoff-Kardjalieff, Lünne, Kalinina, Strauss, Humpf and Studt.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Bachleitner S., Sørensen J. L., Gacek-Matthews A., Sulyok M., Studt L., Strauss J. (2019). Evidence of a demethylase- independent role for the H3K4- specific histone demethylases in Aspergillus nidulans and Fusarium graminearum secondary metabolism. Front. Microbiol. 10:1759. 10.3389/fmicb.2019.01759 - DOI - PMC - PubMed
-
- Bachleitner S., Sulyok M., Sørensen J. L., Strauss J., Studt L. (2021). The H4K20 methyltransferase Kmt5 is involved in secondary metabolism and stress response in phytopathogenic Fusarium species. (in press). - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous