

This is a preprint.
Proximal immune-epithelial progenitor interactions drive chronic tissue sequelae post COVID-19
- PMID: 37745354
- PMCID: PMC10515929
- DOI: 10.1101/2023.09.13.557622
Proximal immune-epithelial progenitor interactions drive chronic tissue sequelae post COVID-19
Abstract
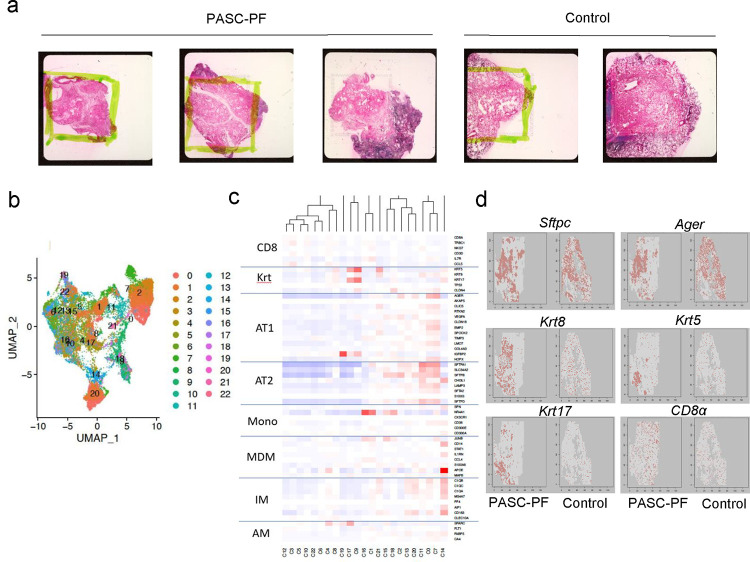
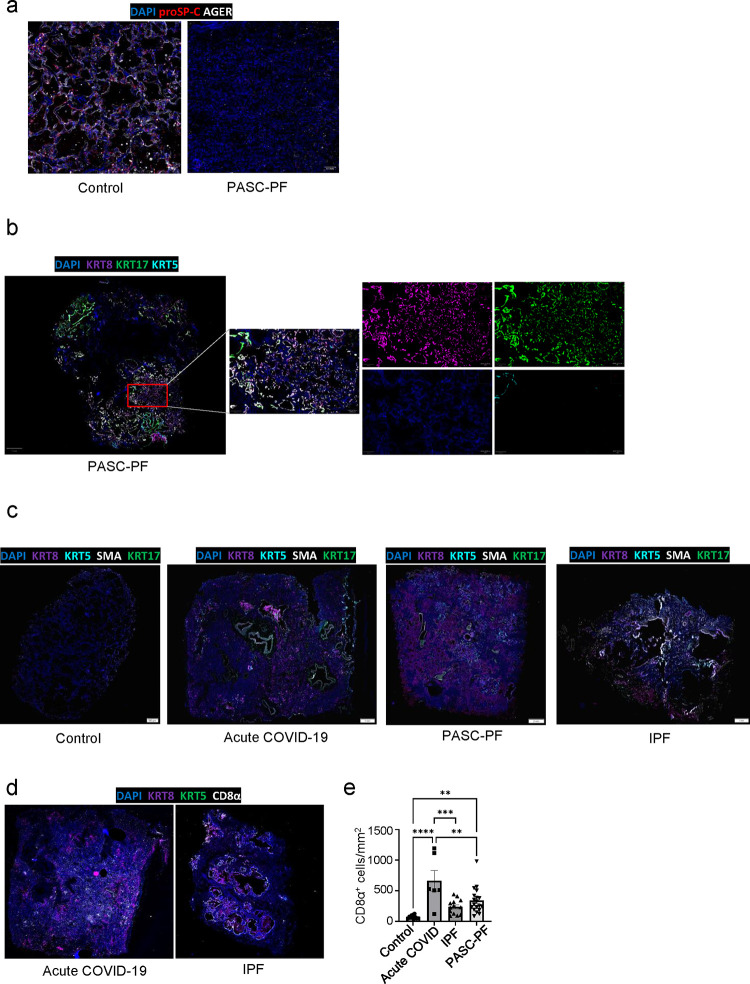
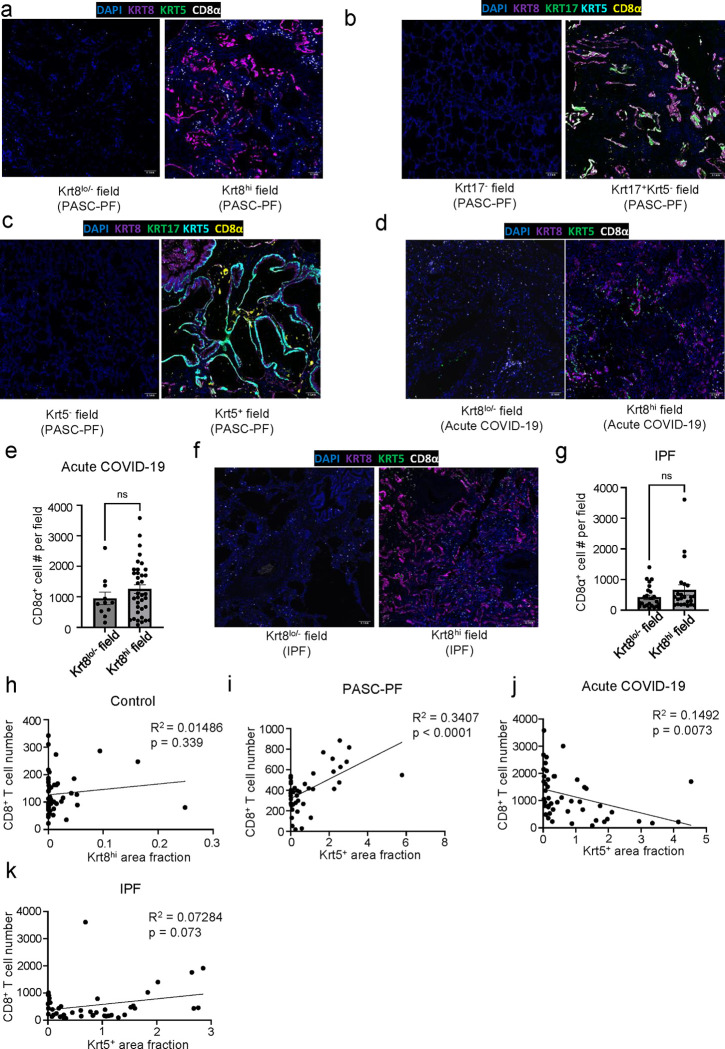
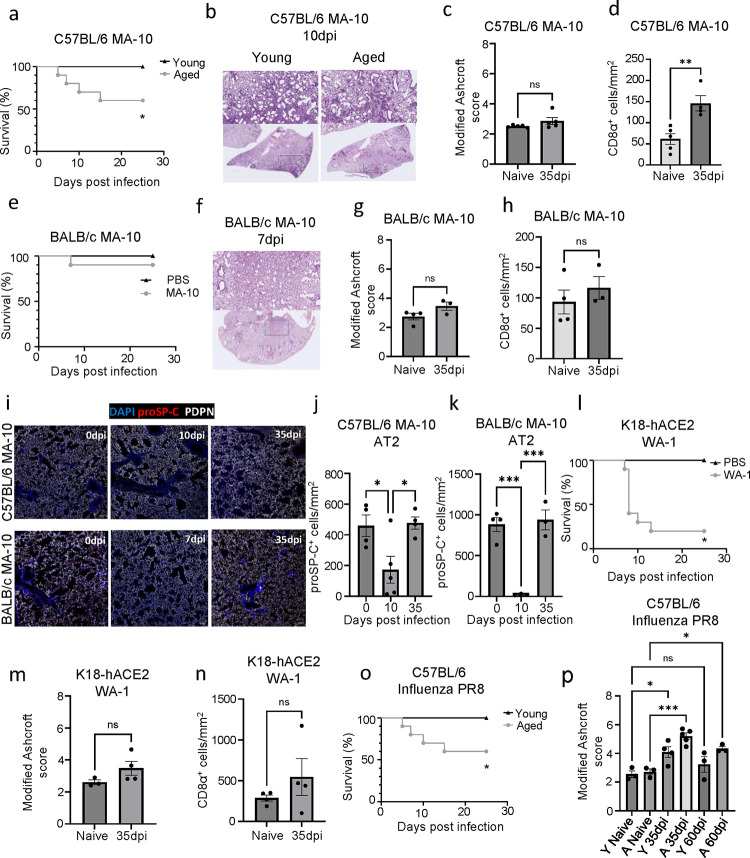
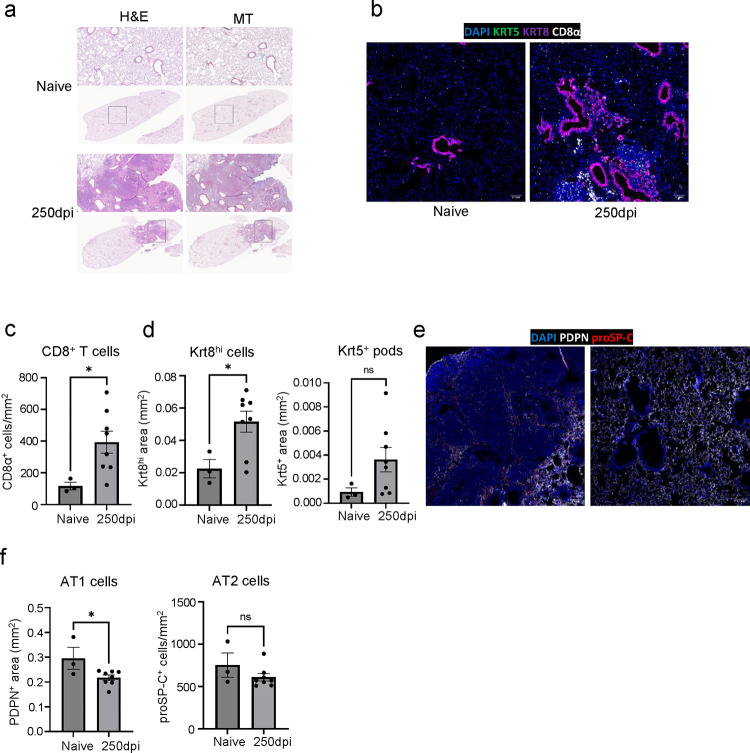
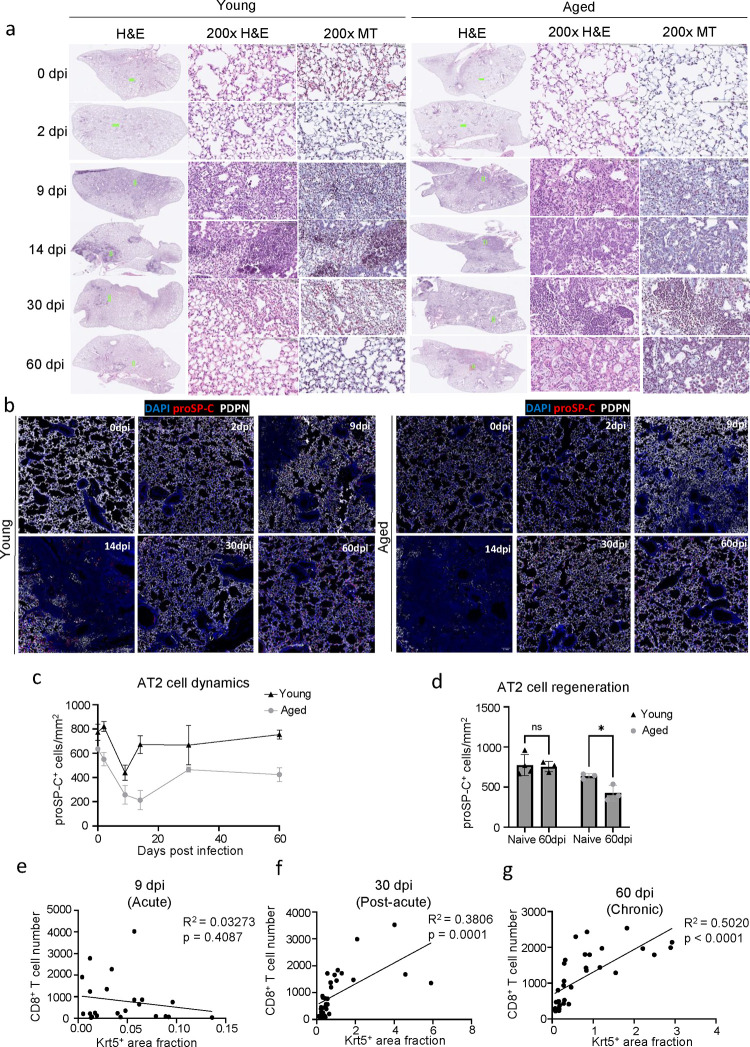
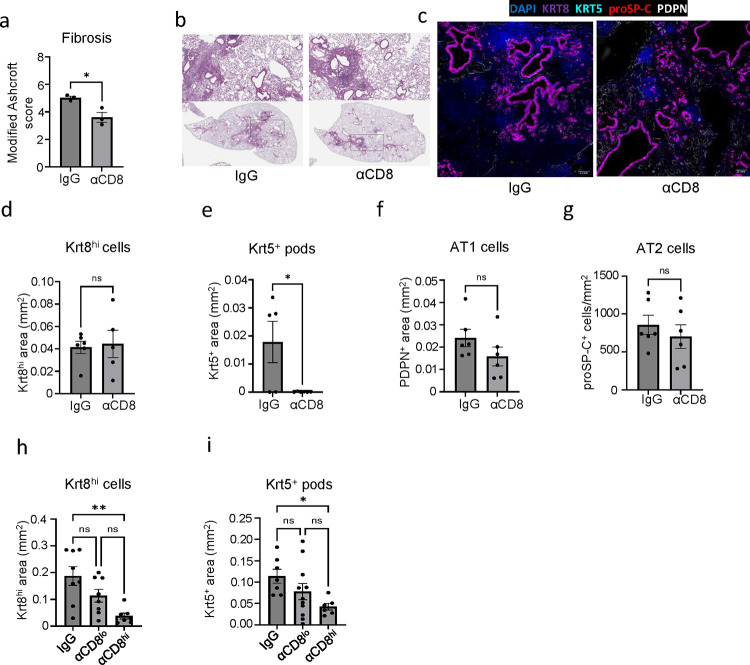
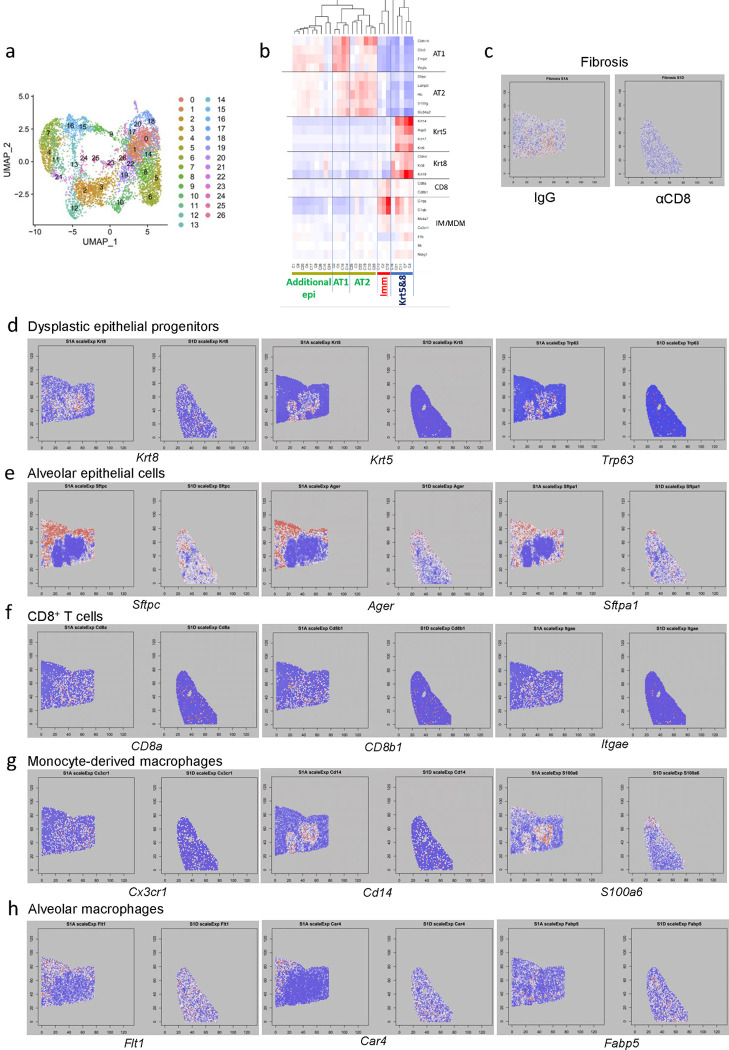
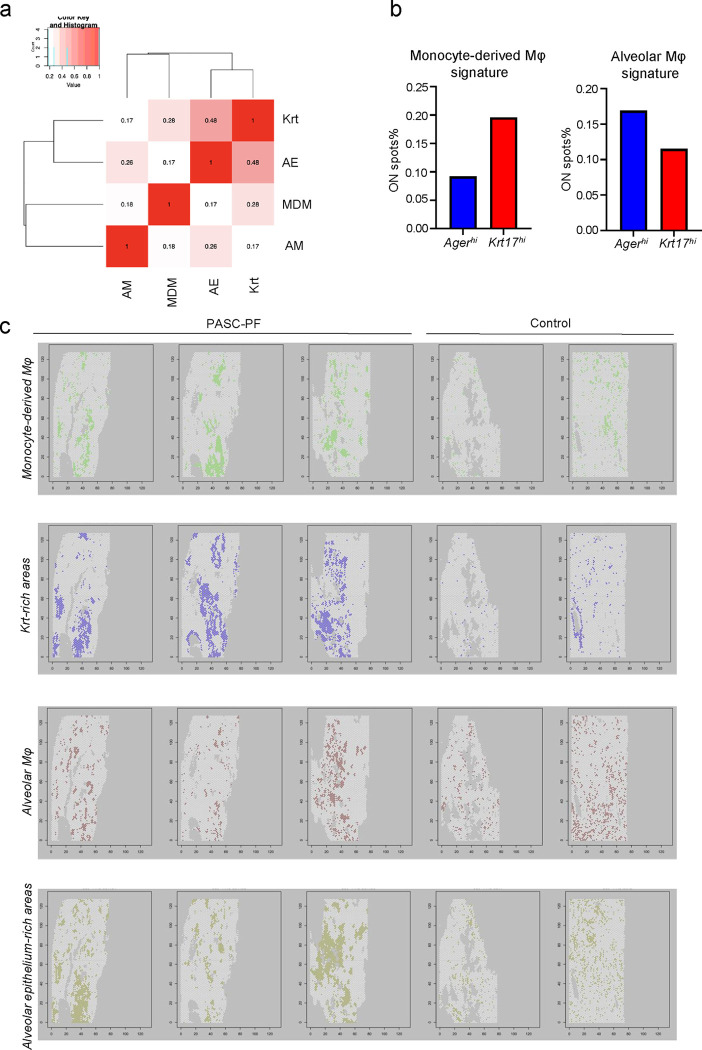
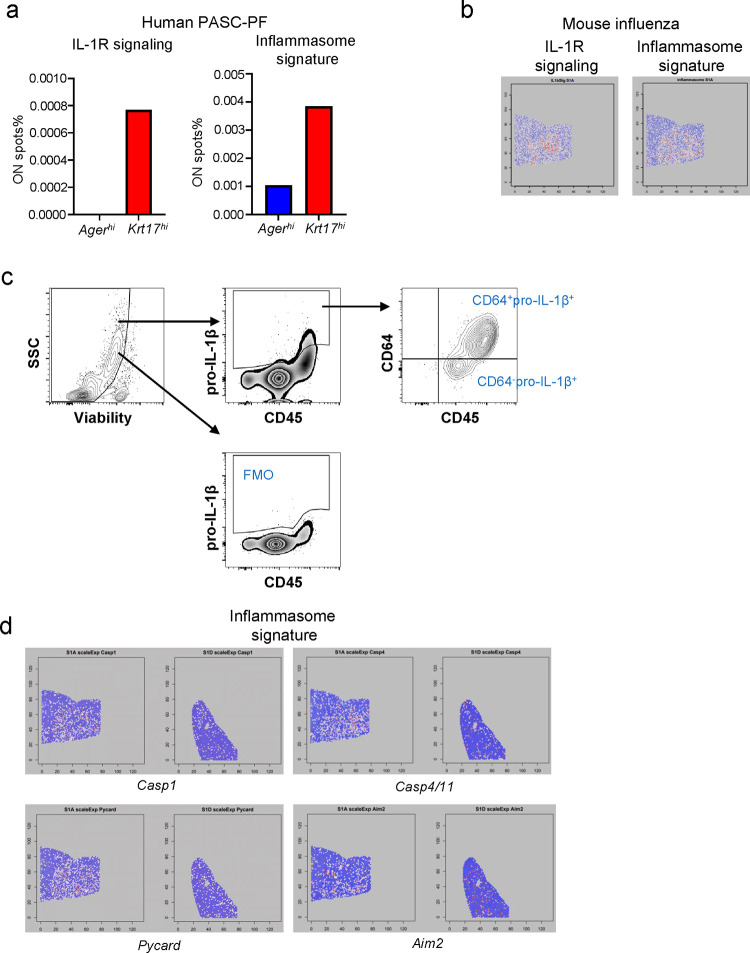
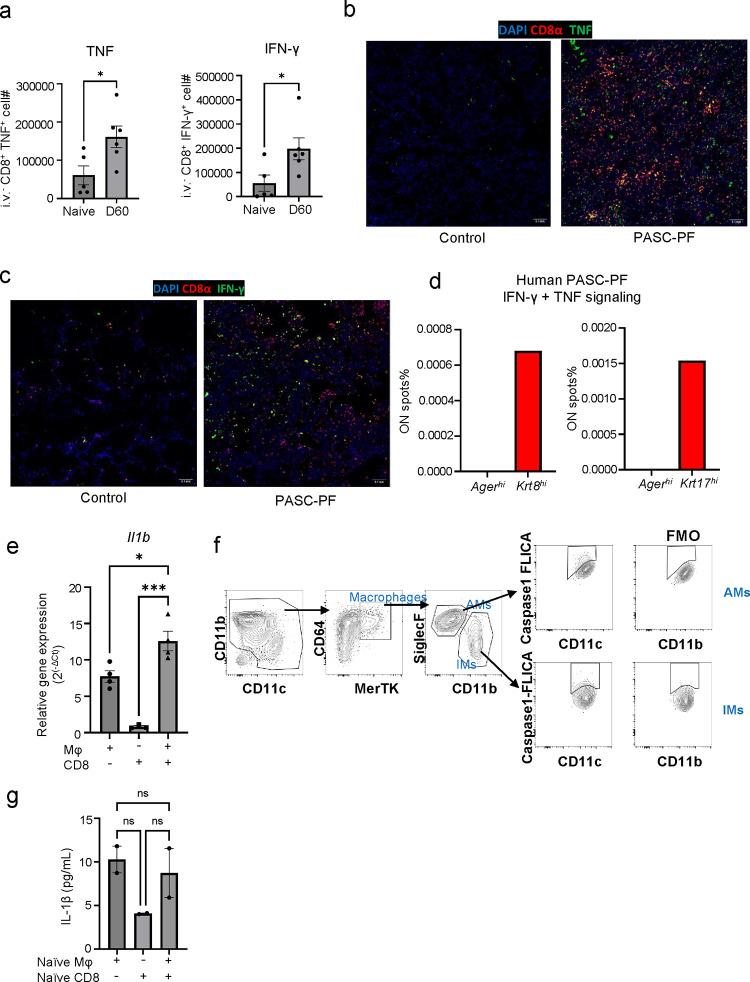
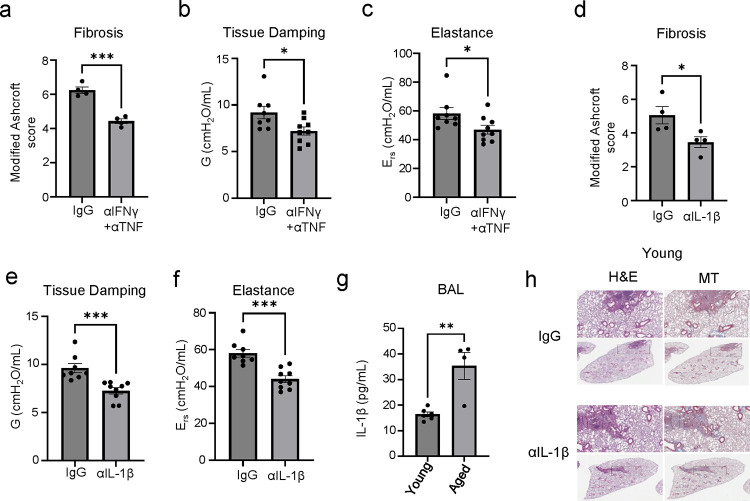
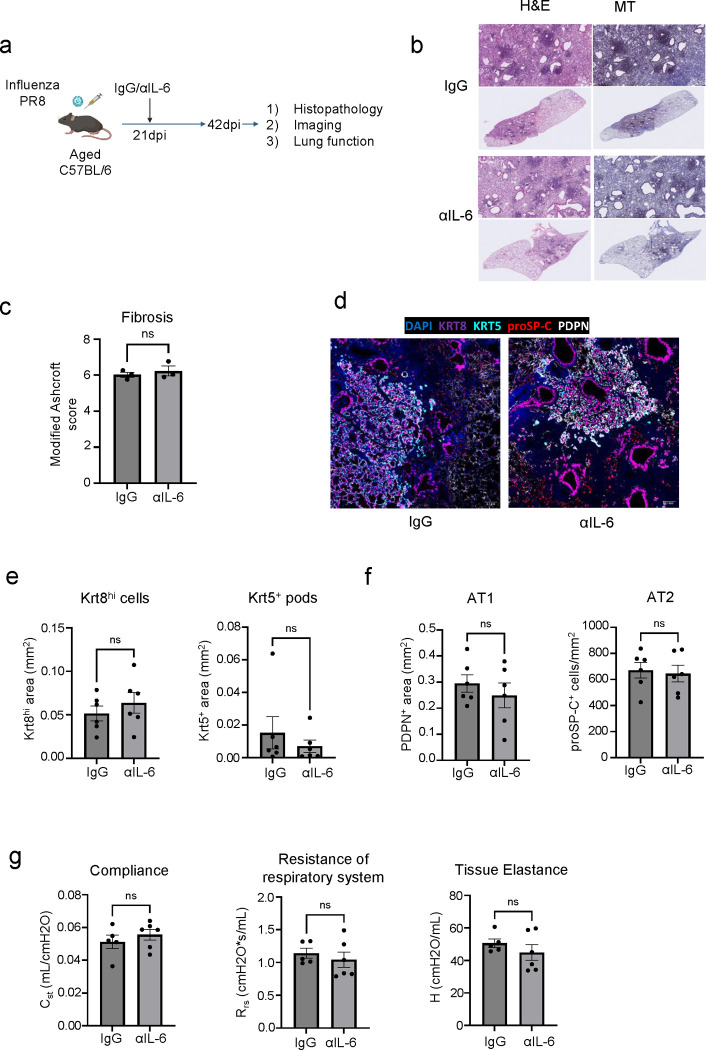
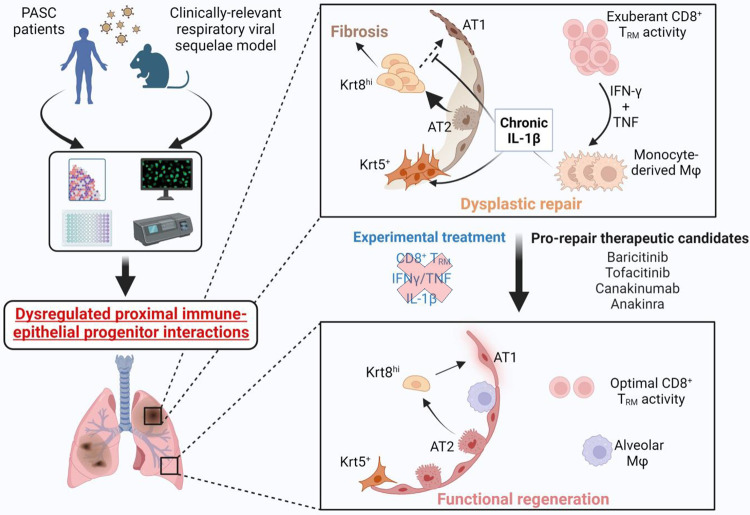
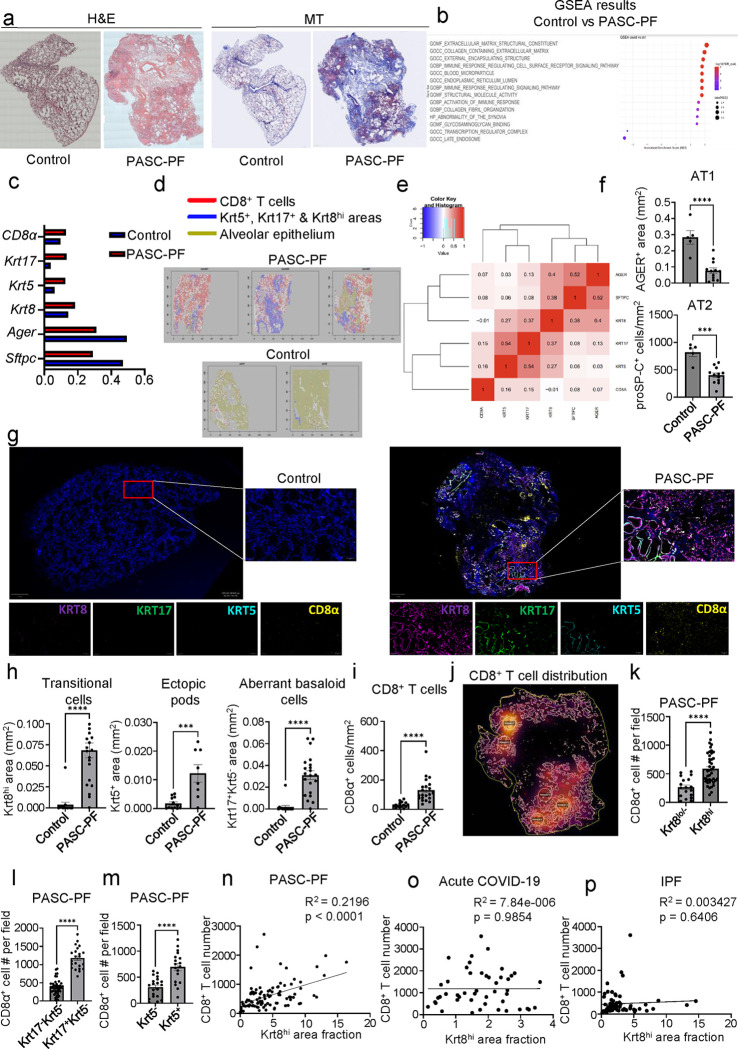
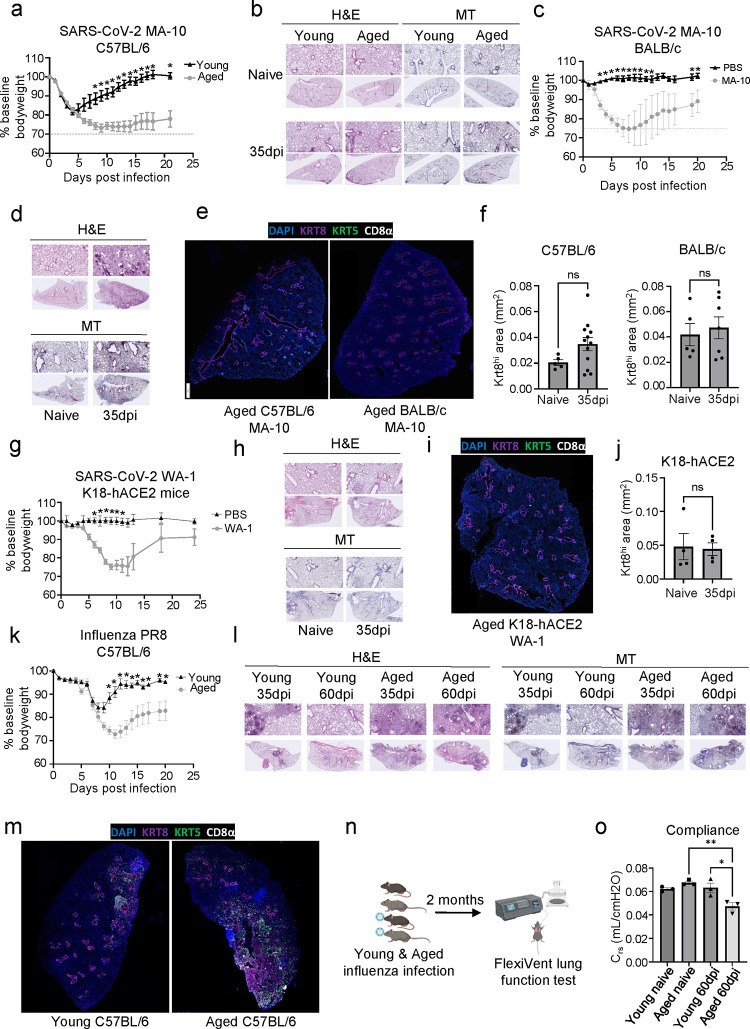
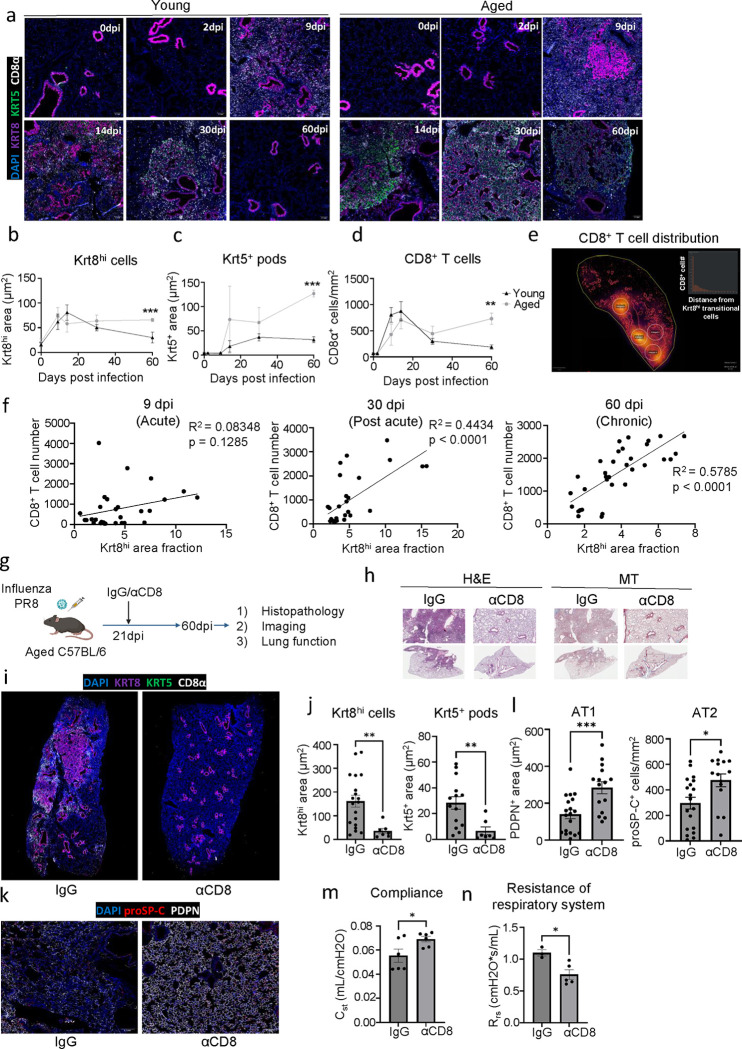
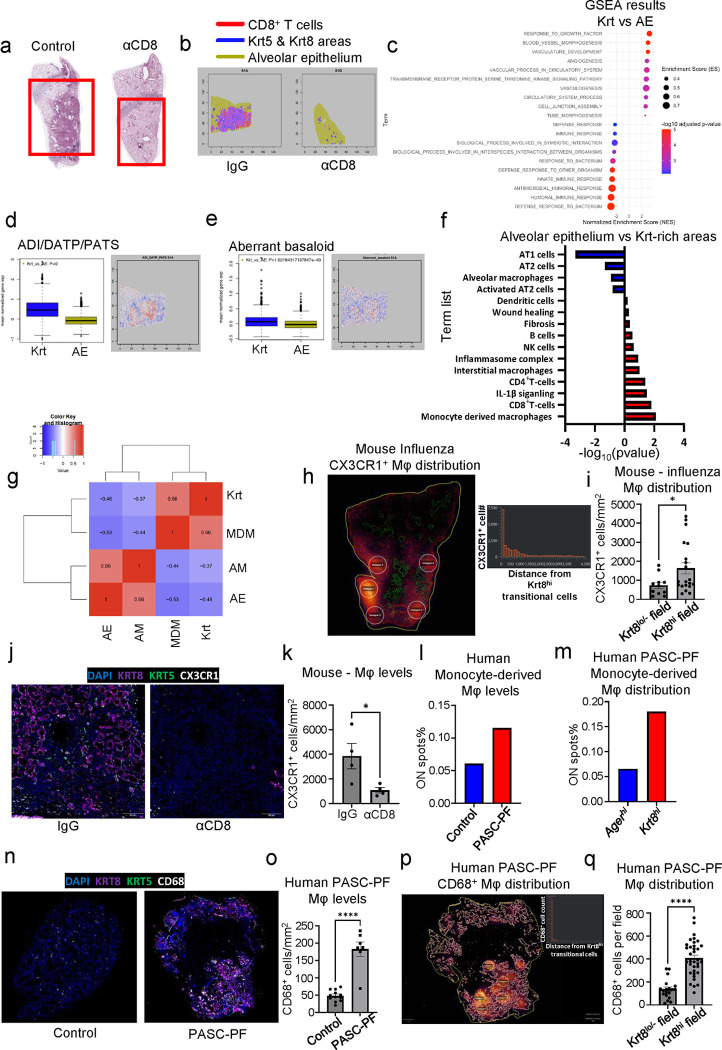
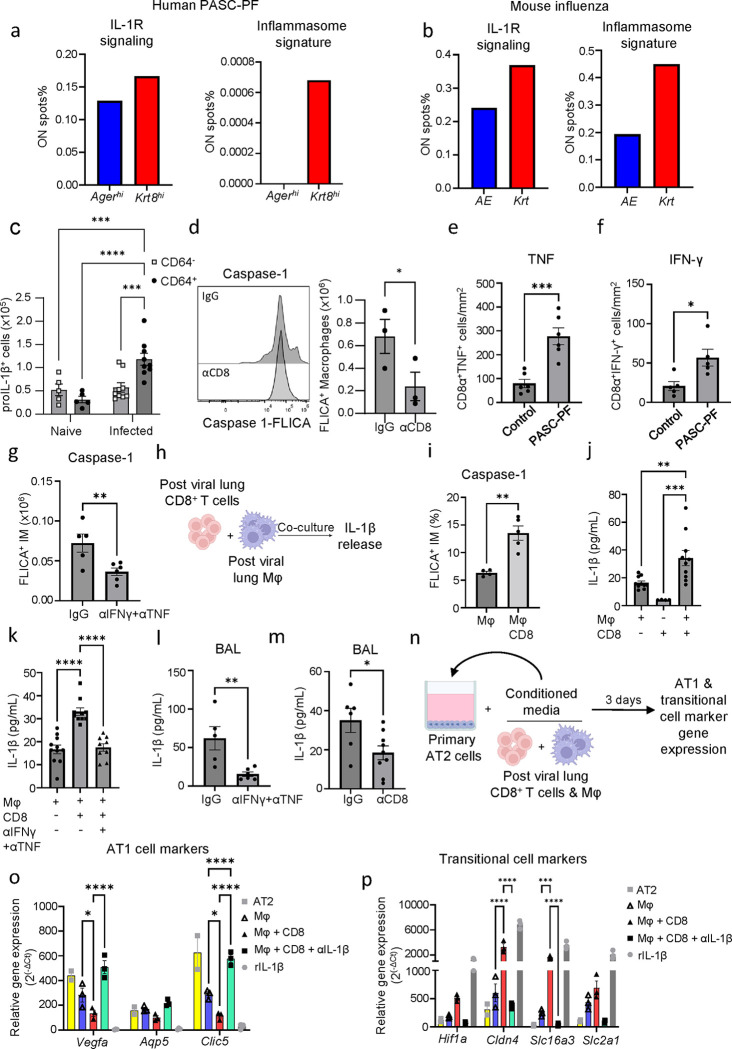
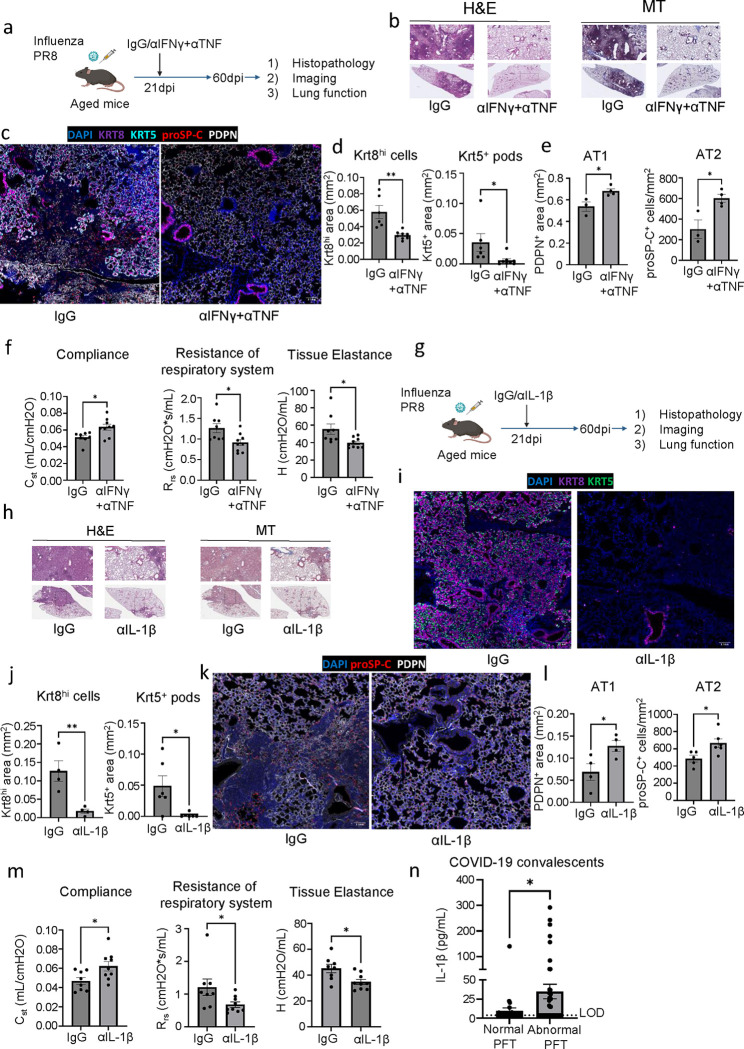
The long-term physiological consequences of SARS-CoV-2, termed Post-Acute Sequelae of COVID-19 (PASC), are rapidly evolving into a major public health concern. The underlying cellular and molecular etiology remain poorly defined but growing evidence links PASC to abnormal immune responses and/or poor organ recovery post-infection. Yet, the precise mechanisms driving non-resolving inflammation and impaired tissue repair in the context of PASC remain unclear. With insights from three independent clinical cohorts of PASC patients with abnormal lung function and/or viral infection-mediated pulmonary fibrosis, we established a clinically relevant mouse model of post-viral lung sequelae to investigate the pathophysiology of respiratory PASC. By employing a combination of spatial transcriptomics and imaging, we identified dysregulated proximal interactions between immune cells and epithelial progenitors unique to the fibroproliferation in respiratory PASC but not acute COVID-19 or idiopathic pulmonary fibrosis (IPF). Specifically, we found a central role for lung-resident CD8+ T cell-macrophage interactions in maintaining Krt8hi transitional and ectopic Krt5+ basal cell progenitors, thus impairing alveolar regeneration and driving fibrotic sequelae after acute viral pneumonia. Mechanistically, CD8+ T cell derived IFN-γ and TNF stimulated lung macrophages to chronically release IL-1β, resulting in the abnormal accumulation of dysplastic epithelial progenitors and fibrosis. Notably, therapeutic neutralization of IFN-γ and TNF, or IL-1β after the resolution of acute infection resulted in markedly improved alveolar regeneration and restoration of pulmonary function. Together, our findings implicate a dysregulated immune-epithelial progenitor niche in driving respiratory PASC. Moreover, in contrast to other approaches requiring early intervention, we highlight therapeutic strategies to rescue fibrotic disease in the aftermath of respiratory viral infections, addressing the current unmet need in the clinical management of PASC and post-viral disease.
Figures
References
-
- Al-Aly Z, Xie Y, Bowe B. High-dimensional characterization of post-acute sequelae of COVID-19. Nature. 2021;594(7862):259–64. - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous