

This is a preprint.
Cerebellar Purkinje Cells Control Posture in Larval Zebrafish (Danio rerio)
- PMID: 37745506
- PMCID: PMC10515840
- DOI: 10.1101/2023.09.12.557469
Cerebellar Purkinje Cells Control Posture in Larval Zebrafish (Danio rerio)
Update in
-
Cerebellar Purkinje cells control posture in larval zebrafish (Danio rerio).Elife. 2025 Apr 24;13:RP97614. doi: 10.7554/eLife.97614. Elife. 2025. PMID: 40272244 Free PMC article.
Abstract
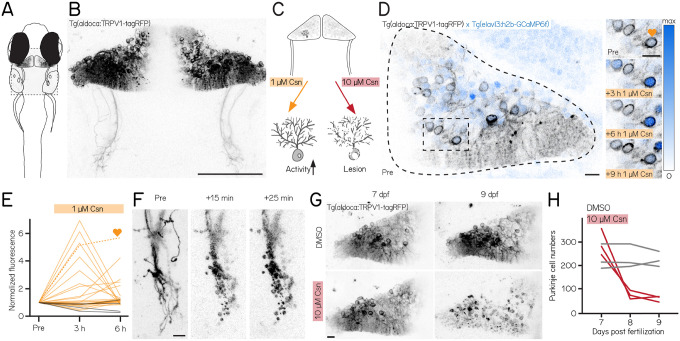
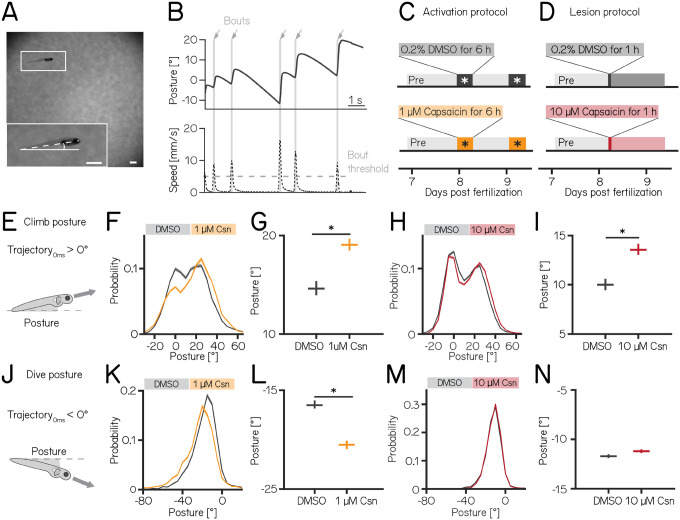
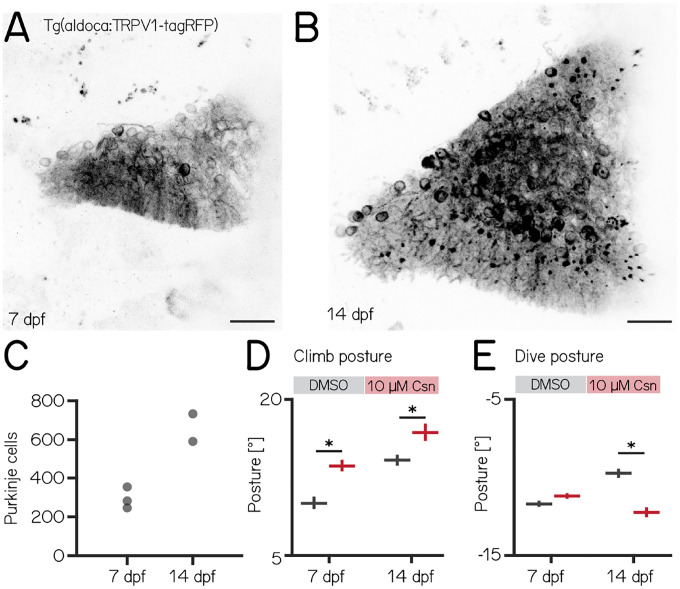
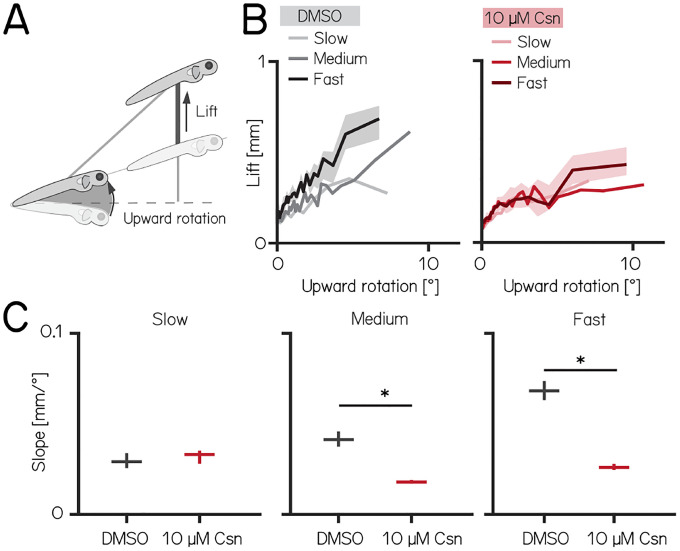
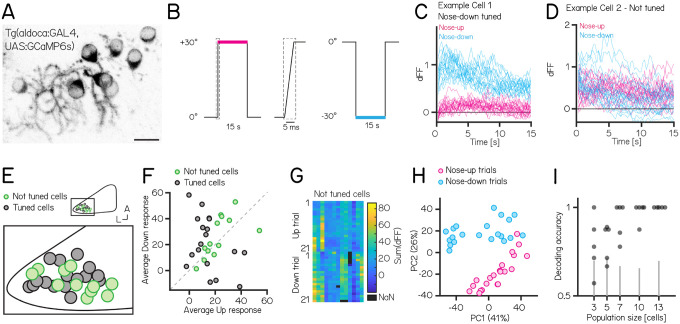
Cerebellar dysfunction leads to postural instability. Recent work in freely moving rodents has transformed investigations of cerebellar contributions to posture. However, the combined complexity of terrestrial locomotion and the rodent cerebellum motivate new approaches to perturb cerebellar function in simpler vertebrates. Here, we adapted a validated chemogenetic tool (TRPV1/capsaicin) to describe the role of Purkinje cells - the output neurons of the cerebellar cortex - as larval zebrafish swam freely in depth. We achieved both bidirectional control (activation and ablation) of Purkinje cells while performing quantitative high-throughput assessment of posture and locomotion. Activation modified postural control in the pitch (nose-up/nose-down) axis. Similarly, ablations disrupted pitch-axis posture and fin-body coordination responsible for climbs. Postural disruption was more widespread in older larvae, offering a window into emergent roles for the developing cerebellum in the control of posture. Finally, we found that activity in Purkinje cells could individually and collectively encode tilt direction, a key feature of postural control neurons. Our findings delineate an expected role for the cerebellum in postural control and vestibular sensation in larval zebrafish, establishing the validity of TRPV1/capsaicin-mediated perturbations in a simple, genetically-tractable vertebrate. Moreover, by comparing the contributions of Purkinje cell ablations to posture in time, we uncover signatures of emerging cerebellar control of posture across early development. This work takes a major step towards understanding an ancestral role of the cerebellum in regulating postural maturation.
Conflict of interest statement
AUTHOR COMPETING INTERESTS The authors declare no competing interests.
Figures
References
-
- Sprague James M. and Chambers William W.. Regulation of posture in intact and decerebrate cat: I. cerebellum, reticular formation, vestibular nuclei. Journal of Neurophysiology, 16(5):451–463, September 1953. - PubMed
-
- Kleine J. F., Guan Y., Kipiani E., Glonti L., Hoshi M., and Büttner U.. Trunk position influences vestibular responses of fastigial nucleus neurons in the alert monkey. Journal of Neurophysiology, 91(5):2090–2100, May 2004. - PubMed
-
- Ioffe M. E.. Cerebellar control of posture. In Handbook of the Cerebellum and Cerebellar Disorders, pages 1221–1240. Springer Netherlands, 2013.
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources