

Anatomical Diversity of the Adult Corticospinal Tract Revealed by Single-Cell Transcriptional Profiling
- PMID: 37748862
- PMCID: PMC10669816
- DOI: 10.1523/JNEUROSCI.0811-22.2023
Anatomical Diversity of the Adult Corticospinal Tract Revealed by Single-Cell Transcriptional Profiling
Abstract
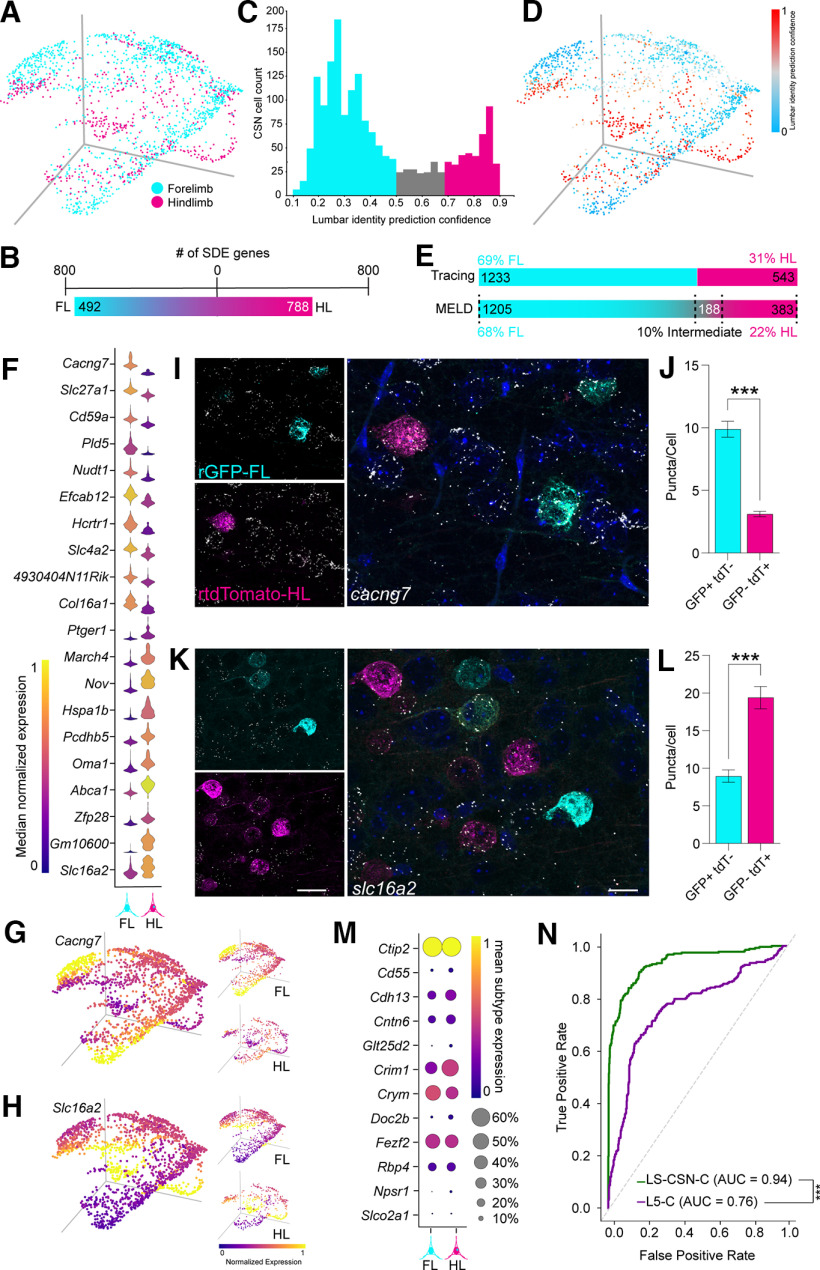
The corticospinal tract (CST) forms a central part of the voluntary motor apparatus in all mammals. Thus, injury, disease, and subsequent degeneration within this pathway result in chronic irreversible functional deficits. Current strategies to repair the damaged CST are suboptimal in part because of underexplored molecular heterogeneity within the adult tract. Here, we combine spinal retrograde CST tracing with single-cell RNA sequencing (scRNAseq) in adult male and female mice to index corticospinal neuron (CSN) subtypes that differentially innervate the forelimb and hindlimb. We exploit publicly available datasets to confer anatomic specialization among CSNs and show that CSNs segregate not only along the forelimb and hindlimb axis but also by supraspinal axon collateralization. These anatomically defined transcriptional data allow us to use machine learning tools to build classifiers that discriminate between CSNs and cortical layer 2/3 and nonspinally terminating layer 5 neurons in M1 and separately identify limb-specific CSNs. Using these tools, CSN subtypes can be differentially identified to study postnatal patterning of the CST in vivo, leveraged to screen for novel limb-specific axon growth survival and growth activators in vitro, and ultimately exploited to repair the damaged CST after injury and disease.SIGNIFICANCE STATEMENT Therapeutic interventions designed to repair the damaged CST after spinal cord injury have remained functionally suboptimal in part because of an incomplete understanding of the molecular heterogeneity among subclasses of CSNs. Here, we combine spinal retrograde labeling with scRNAseq and annotate a CSN index by the termination pattern of their primary axon in the cervical or lumbar spinal cord and supraspinal collateral terminal fields. Using machine learning we have confirmed the veracity of our CSN gene lists to train classifiers to identify CSNs among all classes of neurons in primary motor cortex to study the development, patterning, homeostasis, and response to injury and disease, and ultimately target streamlined repair strategies to this critical motor pathway.
Keywords: Corticospinal tract; plasticity; regeneration; repair; single-cell RNA sequencing; spinal cord injury.
Copyright © 2023 the authors.
Figures






References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases