

Loss of Notch signaling in skeletal stem cells enhances bone formation with aging
- PMID: 37752132
- PMCID: PMC10522593
- DOI: 10.1038/s41413-023-00283-8
Loss of Notch signaling in skeletal stem cells enhances bone formation with aging
Abstract
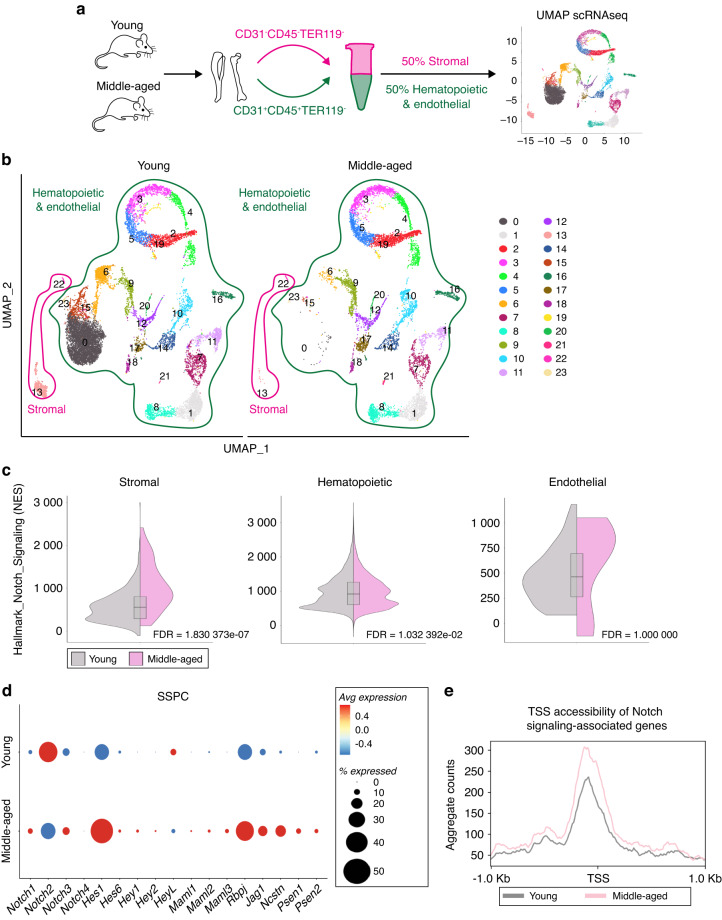
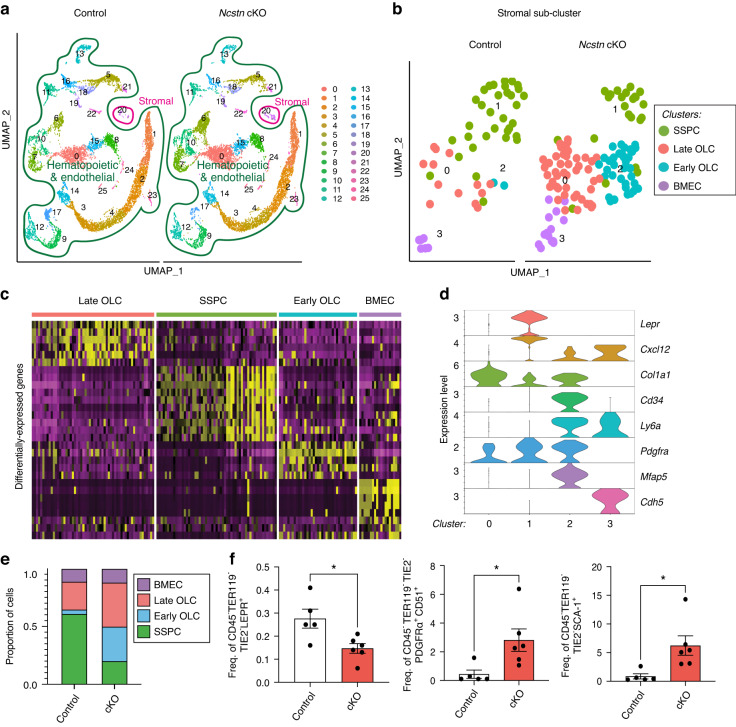
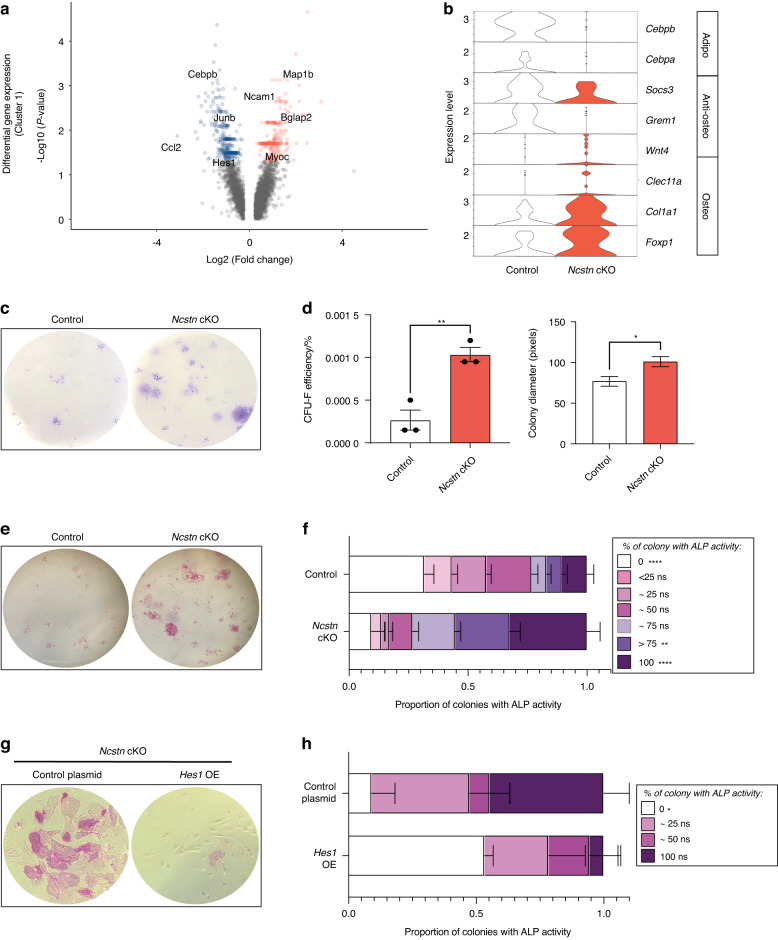
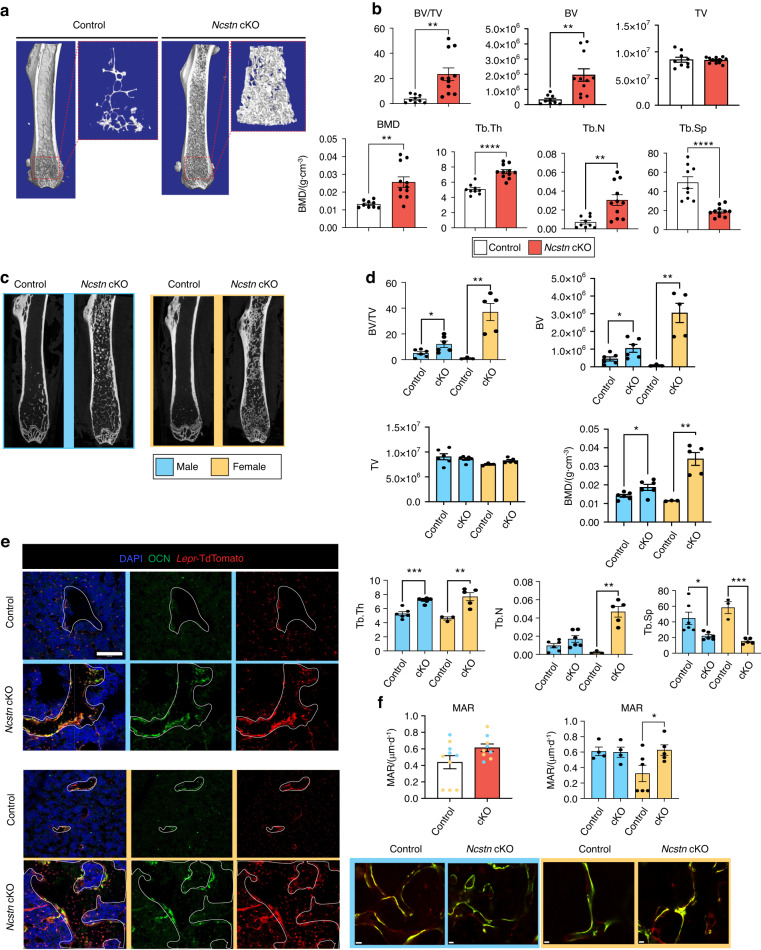
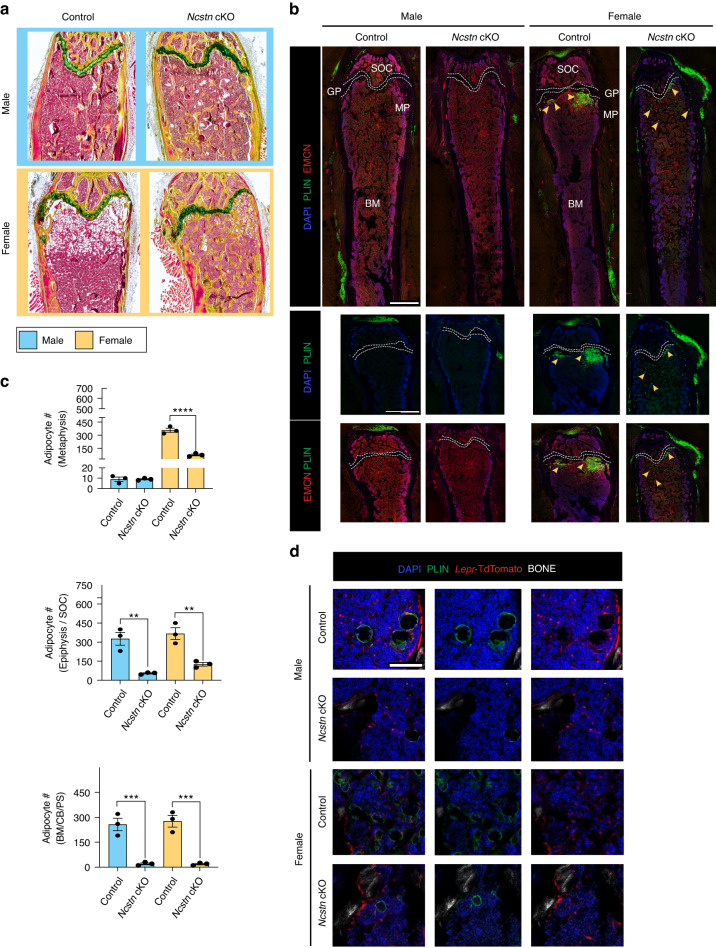
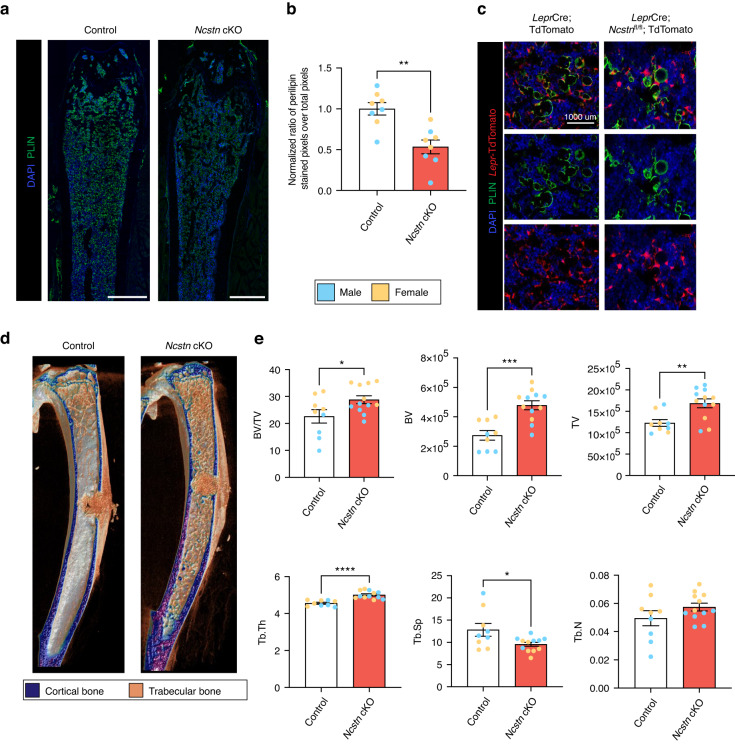
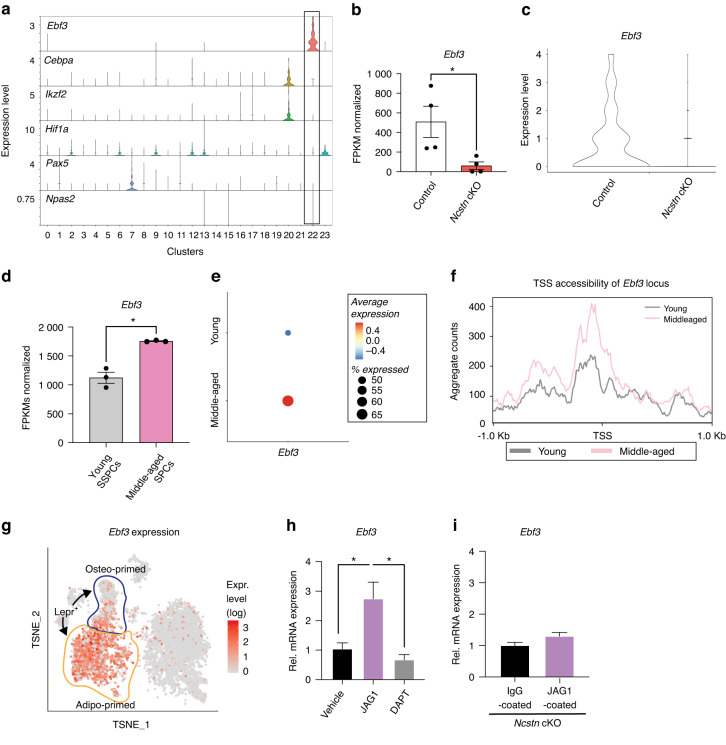
Skeletal stem and progenitor cells (SSPCs) perform bone maintenance and repair. With age, they produce fewer osteoblasts and more adipocytes leading to a loss of skeletal integrity. The molecular mechanisms that underlie this detrimental transformation are largely unknown. Single-cell RNA sequencing revealed that Notch signaling becomes elevated in SSPCs during aging. To examine the role of increased Notch activity, we deleted Nicastrin, an essential Notch pathway component, in SSPCs in vivo. Middle-aged conditional knockout mice displayed elevated SSPC osteo-lineage gene expression, increased trabecular bone mass, reduced bone marrow adiposity, and enhanced bone repair. Thus, Notch regulates SSPC cell fate decisions, and moderating Notch signaling ameliorates the skeletal aging phenotype, increasing bone mass even beyond that of young mice. Finally, we identified the transcription factor Ebf3 as a downstream mediator of Notch signaling in SSPCs that is dysregulated with aging, highlighting it as a promising therapeutic target to rejuvenate the aged skeleton.
© 2023. West China School of Stomatology Sichuan University.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Nishida S, Endo N, Yamagiwa H, Tanizawa T, Takahashi HE. Number of osteoprogenitor cells in human bone marrow markedly decreases after skeletal maturation. J. Bone Miner. Metab. 1999;17:171–177. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
