

Social memory deficit caused by dysregulation of the cerebellar vermis
- PMID: 37752149
- PMCID: PMC10522595
- DOI: 10.1038/s41467-023-41744-2
Social memory deficit caused by dysregulation of the cerebellar vermis
Abstract
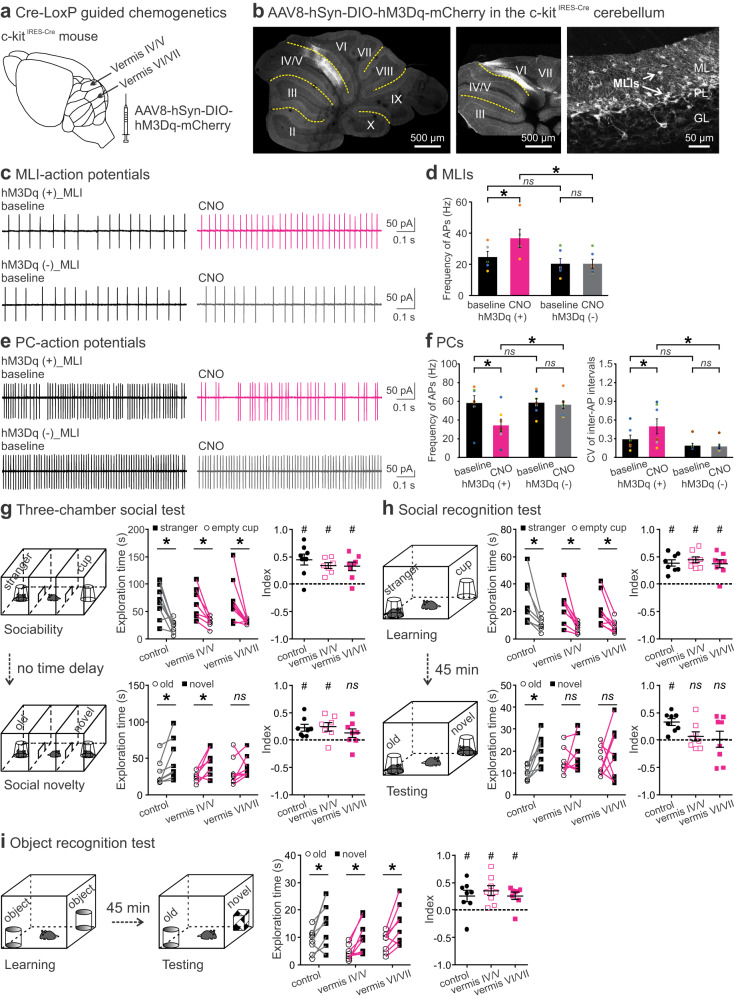
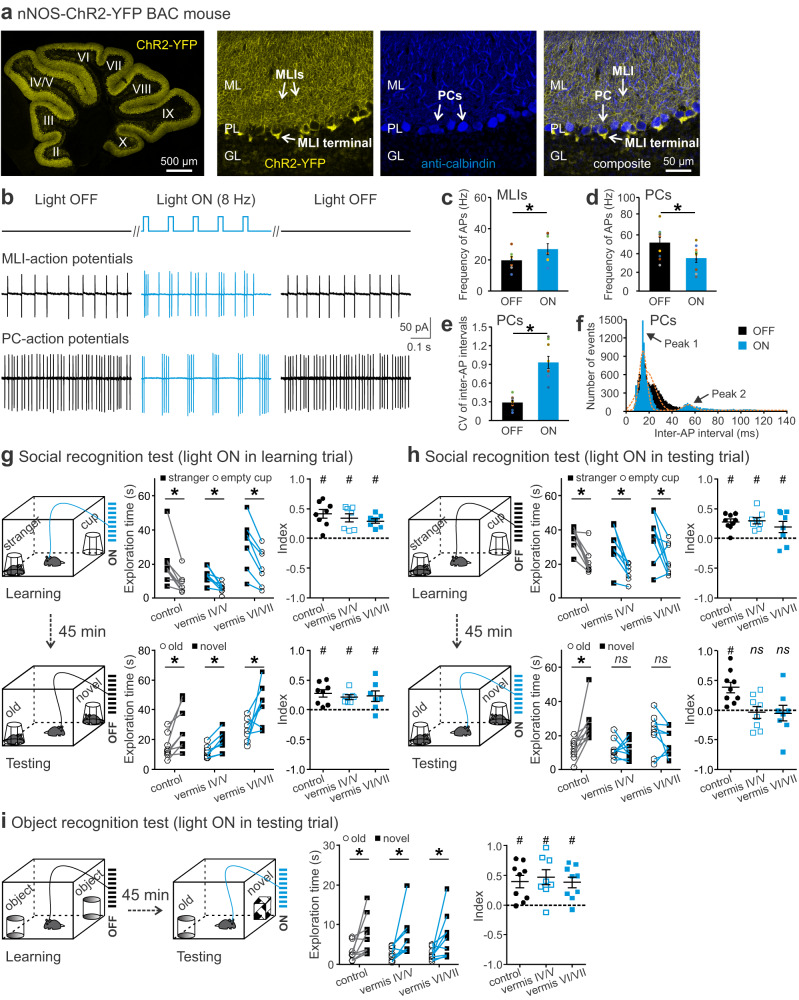
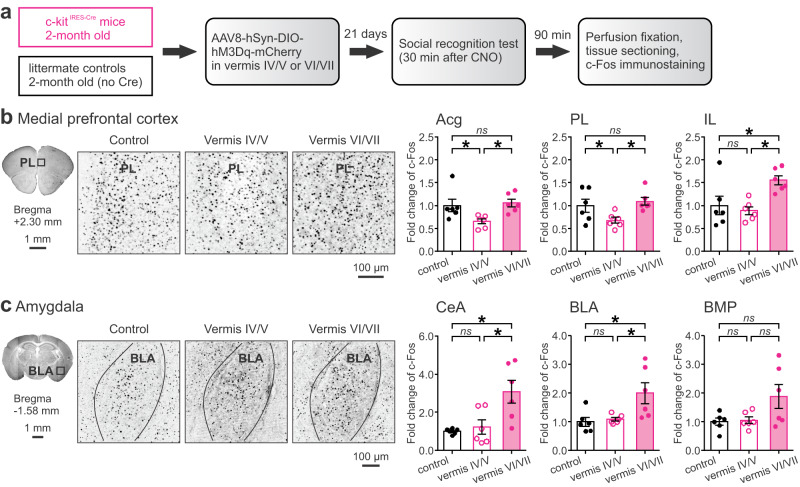
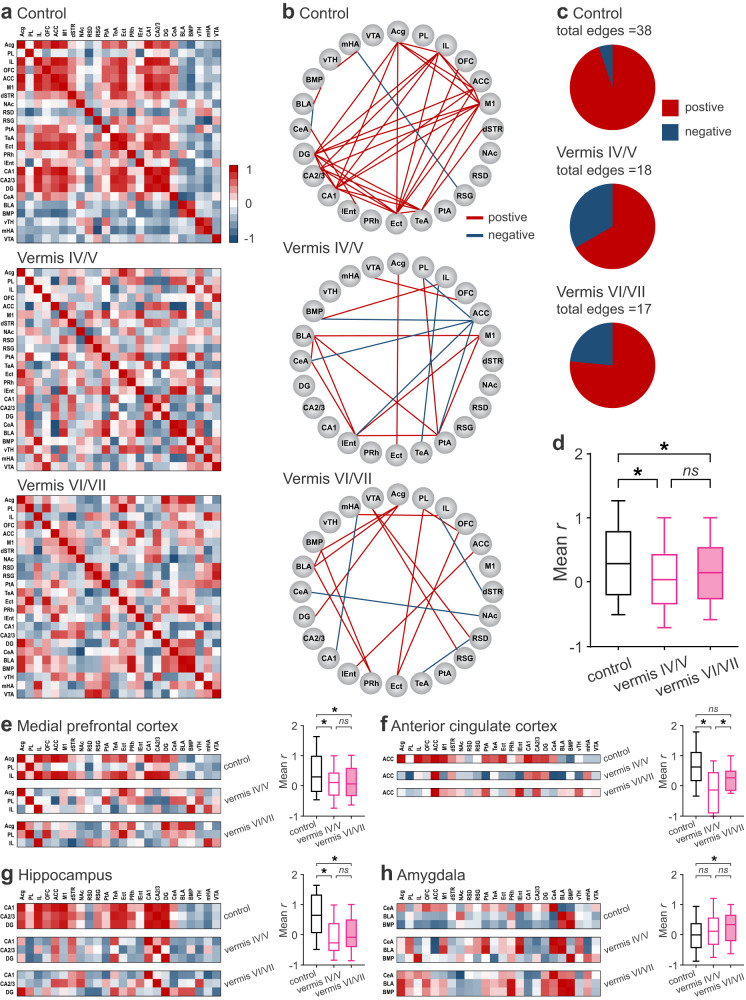
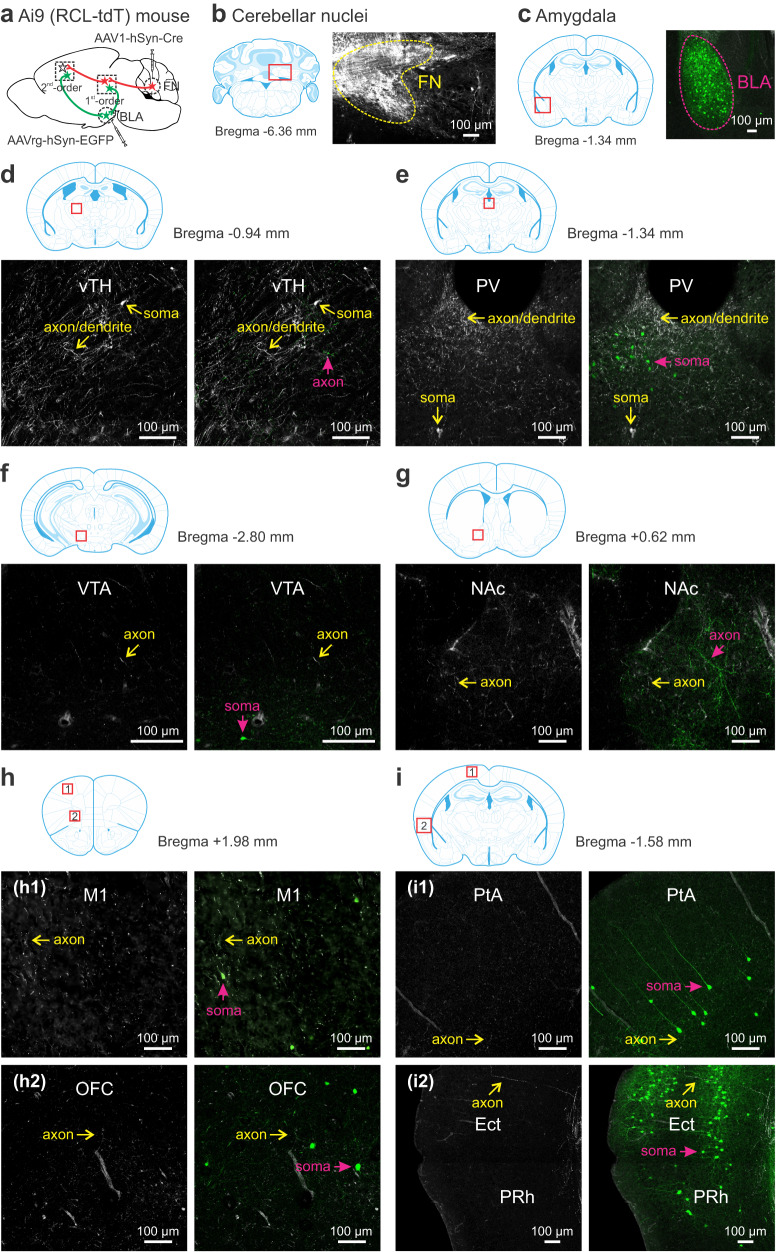
Social recognition memory (SRM) is a key determinant of social interactions. While the cerebellum emerges as an important region for social behavior, how cerebellar activity affects social functions remains unclear. We selectively increased the excitability of molecular layer interneurons (MLIs) to suppress Purkinje cell firing in the mouse cerebellar vermis. Chemogenetic perturbation of MLIs impaired SRM without affecting sociability, anxiety levels, motor coordination or object recognition. Optogenetic interference of MLIs during distinct phases of a social recognition test revealed the cerebellar engagement in the retrieval, but not encoding, of social information. c-Fos mapping after the social recognition test showed that cerebellar manipulation decreased brain-wide interregional correlations and altered network structure from medial prefrontal cortex and hippocampus-centered to amygdala-centered modules. Anatomical tracing demonstrated hierarchical projections from the central cerebellum to the social brain network integrating amygdalar connections. Our findings suggest that the cerebellum organizes the neural matrix necessary for SRM.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
