

Harnessing a mesopelagic predator as a biological sampler reveals taxonomic and vertical resource partitioning among three poorly known deep-sea fishes
- PMID: 37752192
- PMCID: PMC10522621
- DOI: 10.1038/s41598-023-41298-9
Harnessing a mesopelagic predator as a biological sampler reveals taxonomic and vertical resource partitioning among three poorly known deep-sea fishes
Abstract
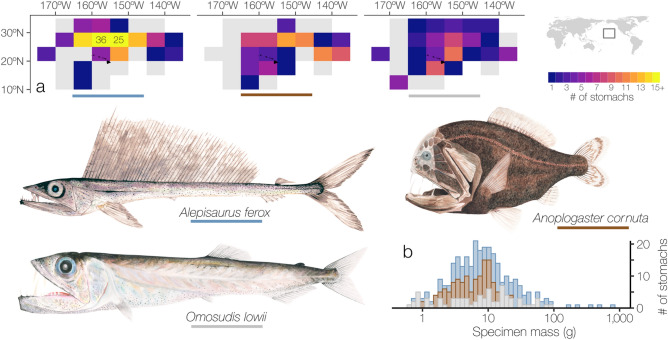
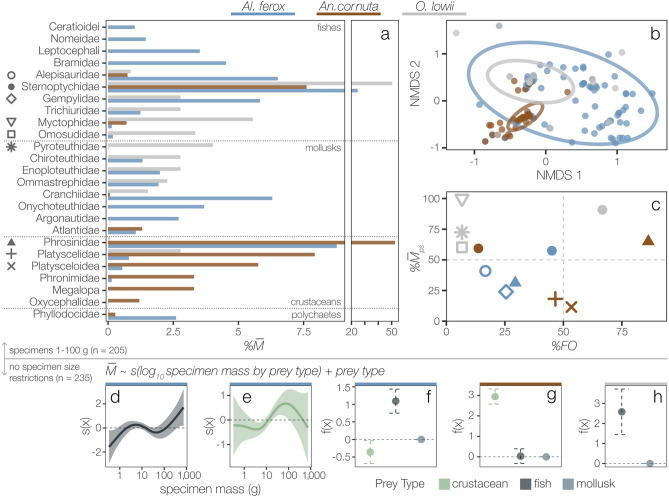
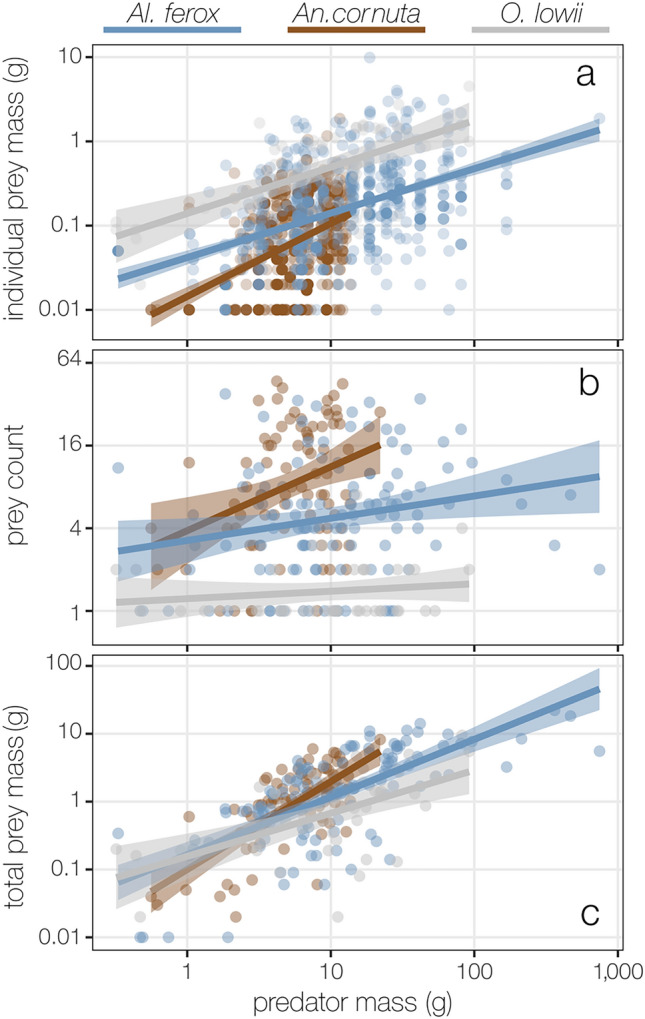
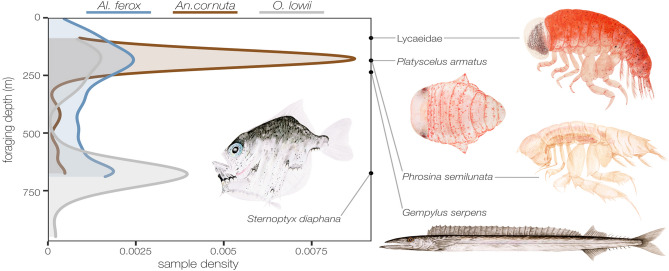
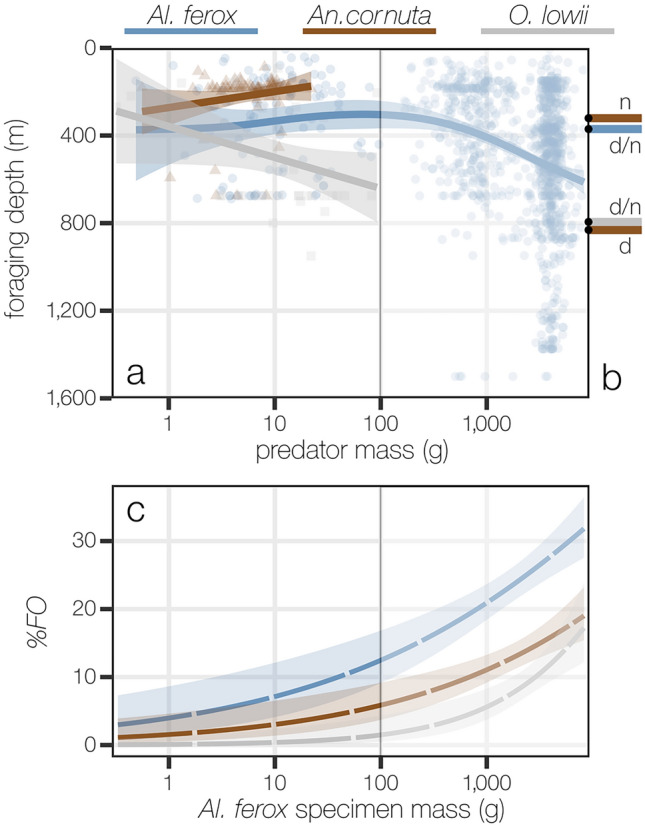
Pelagic predators are effective biological samplers of midtrophic taxa and are especially useful in deep-sea habitats where relatively mobile taxa frequently avoid observation with conventional methods. We examined specimens sampled from the stomachs of longnose lancetfish, Alepisaurus ferox, to describe the diets and foraging behaviors of three common, but poorly known deep-sea fishes: the hammerjaw (Omosudis lowii, n = 79, 0.3-92 g), juvenile common fangtooth (Anoplogaster cornuta, n = 91, 0.6-22 g), and juvenile Al. ferox (n = 138, 0.3-744 g). Diet overlap among the three species was high, with five shared prey families accounting for 63 ± 11% of the total prey mass per species. However, distinct differences in foraging strategies and prey sizes were evident. Resource partitioning was greatest between An. cornuta that specialized on small (mean = 0.13 ± 0.11 g), shallow-living hyperiid amphipods and O. lowii that specialized on large (mean = 0.97 ± 0.45 g), deep-dwelling hatchetfishes. Juvenile Al. ferox foraged on a high diversity of prey from both shallow and deep habitats. We describe the foraging ecologies of three midtrophic fish competitors and demonstrate the potential for biological samplers to improve our understanding of deep-sea food webs.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Gastauer S, Nickels CF, Ohman MD. Body size- and season-dependent diel vertical migration of mesozooplankton resolved acoustically in the San Diego Trough. Limnol. Oceanogr. 2022;67:300–313. doi: 10.1002/lno.11993. - DOI
-
- Staby A, Srisomwong J, Rosland R. Variation in DVM behaviour of juvenile and adult pearlside (Maurolicus muelleri) linked to feeding strategies and related predation risk. Fish. Oceanogr. 2013;22:90–101. doi: 10.1111/fog.12012. - DOI
