

Contribution of viral and bacterial infections to senescence and immunosenescence
- PMID: 37753486
- PMCID: PMC10518457
- DOI: 10.3389/fcimb.2023.1229098
Contribution of viral and bacterial infections to senescence and immunosenescence
Abstract
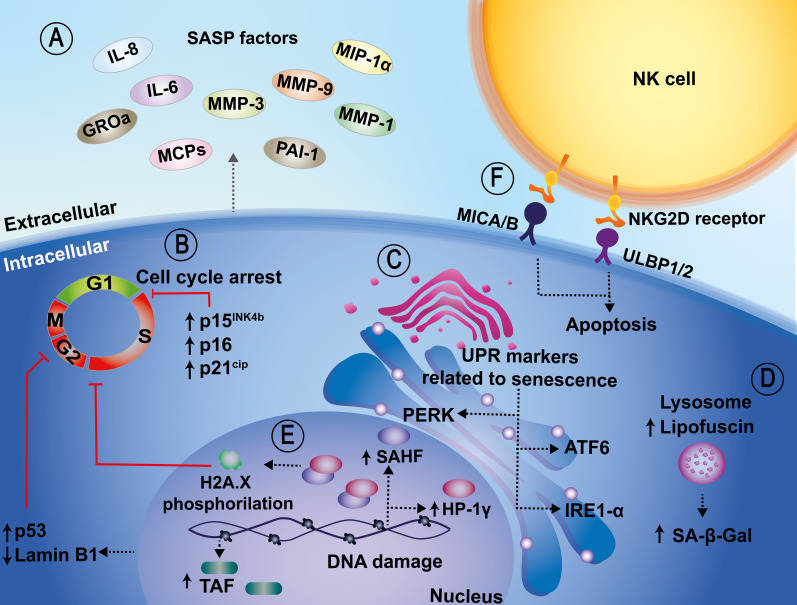
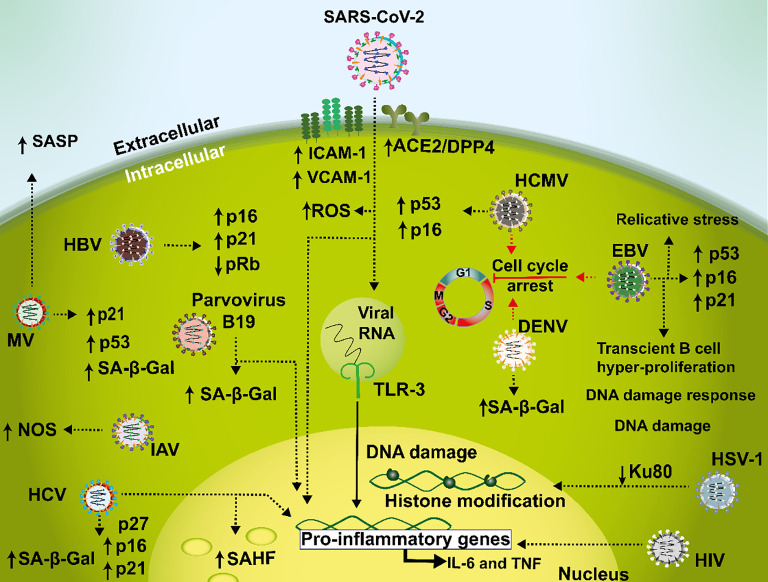
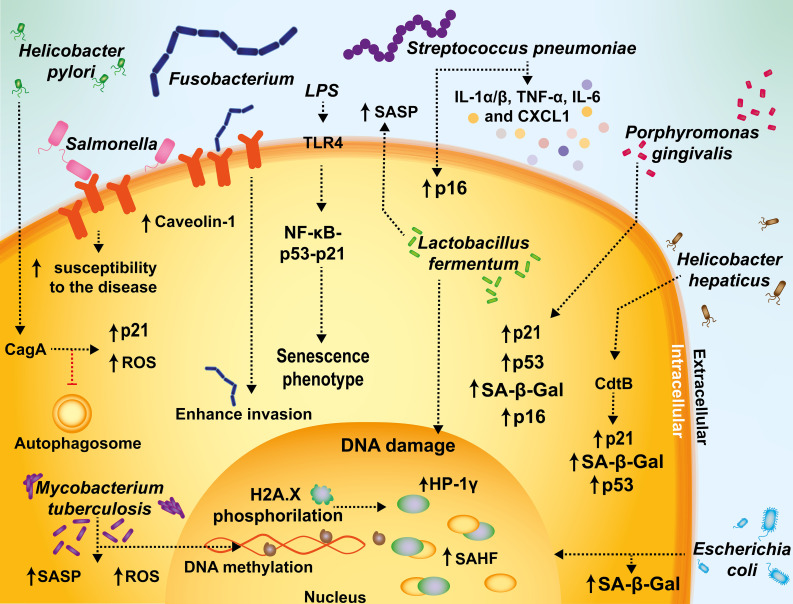
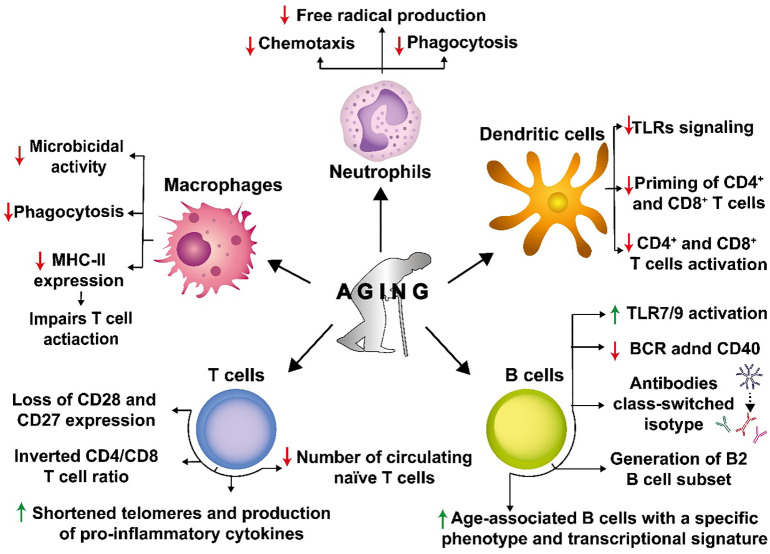
Cellular senescence is a key biological process characterized by irreversible cell cycle arrest. The accumulation of senescent cells creates a pro-inflammatory environment that can negatively affect tissue functions and may promote the development of aging-related diseases. Typical biomarkers related to senescence include senescence-associated β-galactosidase activity, histone H2A.X phosphorylation at serine139 (γH2A.X), and senescence-associated heterochromatin foci (SAHF) with heterochromatin protein 1γ (HP-1γ protein) Moreover, immune cells undergoing senescence, which is known as immunosenescence, can affect innate and adaptative immune functions and may elicit detrimental effects over the host's susceptibility to infectious diseases. Although associations between senescence and pathogens have been reported, clear links between both, and the related molecular mechanisms involved remain to be determined. Furthermore, it remains to be determined whether infections effectively induce senescence, the impact of senescence and immunosenescence over infections, or if both events coincidently share common molecular markers, such as γH2A.X and p53. Here, we review and discuss the most recent reports that describe cellular hallmarks and biomarkers related to senescence in immune and non-immune cells in the context of infections, seeking to better understand their relationships. Related literature was searched in Pubmed and Google Scholar databases with search terms related to the sections and subsections of this review.
Keywords: SASP; bacteria; chronic infections; immunosenescence; persistent infections; senescence; virus.
Copyright © 2023 Reyes, Ortiz, Duarte, Fernández, Hernández-Armengol, Palacios, Prado, Andrade, Rodriguez-Guilarte, Kalergis, Simon, Carreño, Riedel, Cáceres and González.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous