

A global-temporal analysis on Phytophthora sojae resistance-gene efficacy
- PMID: 37758723
- PMCID: PMC10533513
- DOI: 10.1038/s41467-023-41321-7
A global-temporal analysis on Phytophthora sojae resistance-gene efficacy
Abstract
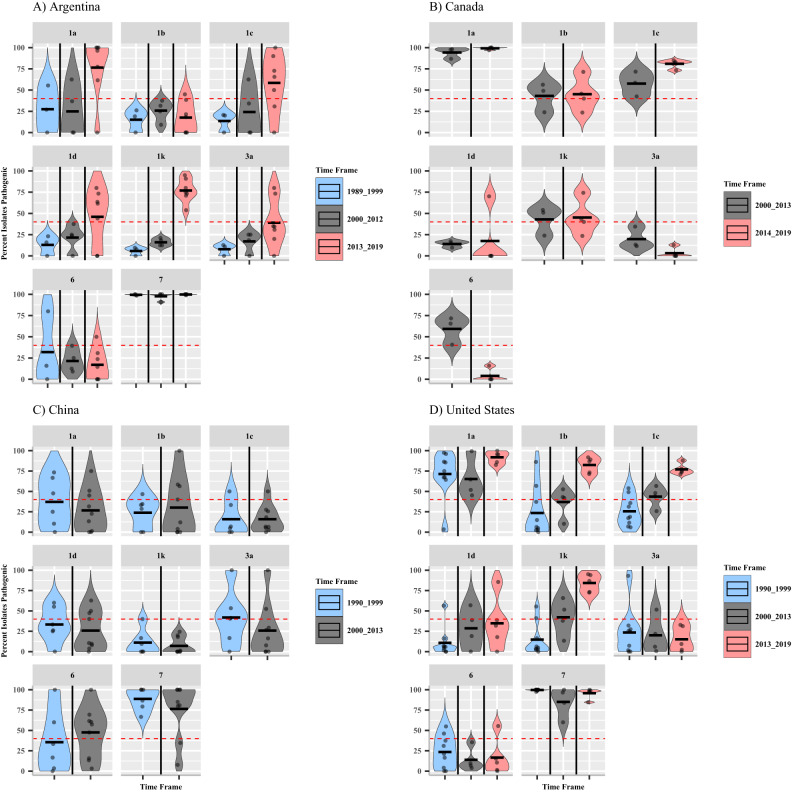
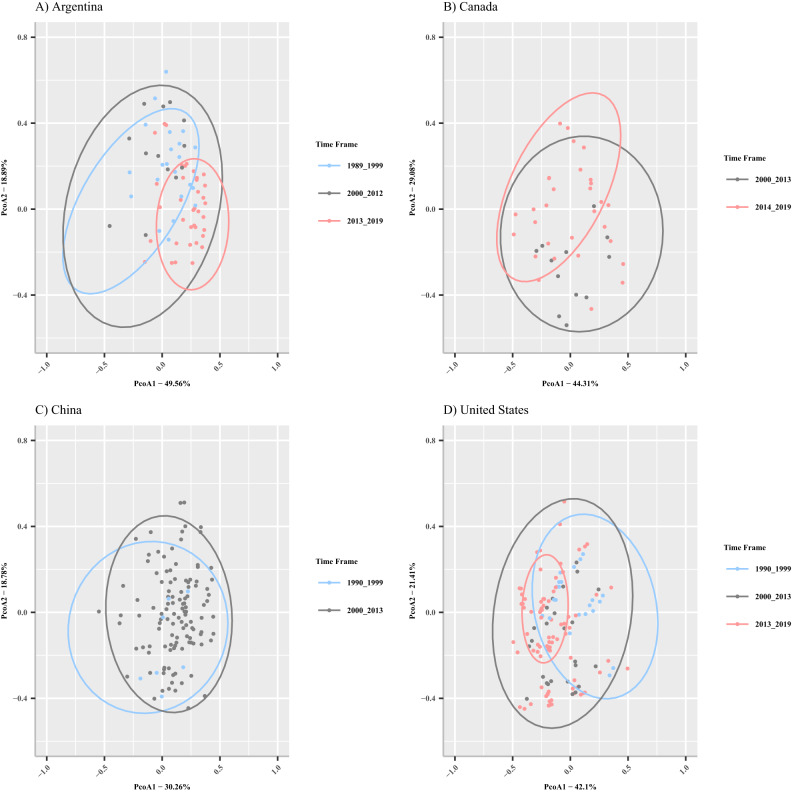
Plant disease resistance genes are widely used in agriculture to reduce disease outbreaks and epidemics and ensure global food security. In soybean, Rps (Resistance to Phytophthora sojae) genes are used to manage Phytophthora sojae, a major oomycete pathogen that causes Phytophthora stem and root rot (PRR) worldwide. This study aims to identify temporal changes in P. sojae pathotype complexity, diversity, and Rps gene efficacy. Pathotype data was collected from 5121 isolates of P. sojae, derived from 29 surveys conducted between 1990 and 2019 across the United States, Argentina, Canada, and China. This systematic review shows a loss of efficacy of specific Rps genes utilized for disease management and a significant increase in the pathotype diversity of isolates over time. This study finds that the most widely deployed Rps genes used to manage PRR globally, Rps1a, Rps1c and Rps1k, are no longer effective for PRR management in the United States, Argentina, and Canada. This systematic review emphasizes the need to widely introduce new sources of resistance to P. sojae, such as Rps3a, Rps6, or Rps11, into commercial cultivars to effectively manage PRR going forward.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures

References
-
- United States Department of Agriculture Foreign Agriculture Service. World Agricultural Production. Circular Series WAP 8-22, pp. 1–47 https://apps.fas.usda.gov/psdonline/app/index.html#/app/downloads (Department of Agriculture, Washington, 2022).
-
- United Nations. World Population Projected to Reach 9.8 Billion in 2050, and 11.2 Billion in 2100. https://www.un.org/en/desa/world-population-projected-reach-98-billion-2... (UN, 2017).
-
- Ning Y, Liu W, Wang GL. Balancing immunity and yield in crop plants. Trends Plant Sci. 2017;22:1069–1079. - PubMed
-
- Tyler BM. Phytophthora sojae: root rot pathogen of soybean and model oomycete. Mol. Plant Pathol. 2007;8:1–8. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
