

Biomarker Signature in Aqueous Humor Mirrors Lens Epithelial Cell Activation: New Biomolecular Aspects from Cataractogenic Myopia
- PMID: 37759728
- PMCID: PMC10526747
- DOI: 10.3390/biom13091328
Biomarker Signature in Aqueous Humor Mirrors Lens Epithelial Cell Activation: New Biomolecular Aspects from Cataractogenic Myopia
Abstract
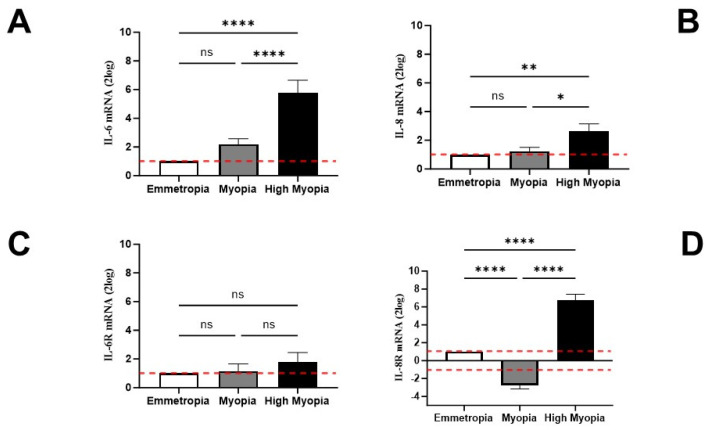
Inflammatory, vasculogenic, and profibrogenic factors have been previously reported in vitreous (VH) and aqueous (AH) humors in myopic patients who underwent cataract surgery. In light of this, we selected some mediators for AH and anterior-capsule-bearing lens epithelial cell (AC/LEC) analysis, and AH expression was correlated with LEC activation (epithelial-mesenchymal transition and EMT differentiation) and axial length (AL) elongation. In this study, AH (97; 41M/56F) and AC/LEC samples (78; 35M/43F) were collected from 102 patients who underwent surgery, and biosamples were grouped according to AL elongation. Biomolecular analyses were carried out for AH and LECs, while microscopical analyses were restricted to whole flattened AC/LECs. The results showed increased levels of interleukin (IL)-6, IL-8, and angiopoietin-2 (ANG)-2 and decreased levels of vascular endothelium growth factor (VEGF)-A were detected in AH depending on AL elongation. LECs showed EMT differentiation as confirmed by the expression of smooth muscle actin (α-SMA) and transforming growth factor (TGF)-βR1/TGFβ isoforms. A differential expression of IL-6R/IL-6, IL-8R/IL-8, and VEGF-R1/VEGF was observed in the LECs, and this expression correlated with AL elongation. The higher VEGF-A and lower VEGF-D transcript expressions were detected in highly myopic LECs, while no significant changes were monitored for VEGF-R transcripts. In conclusion, these findings provide a strong link between the AH protein signature and the EMT phenotype. Furthermore, the low VEGF-A/ANG-2 and the high VEGF-A/VEGF-D ratios in myopic AH might suggest a specific inflammatory and profibrogenic pattern in high myopia. The highly myopic AH profile might be a potential candidate for rating anterior chamber inflammation and predicting retinal distress at the time of cataract surgery.
Keywords: ETM; aqueous; biomarkers; cataract; inflammation; lens epithelial cells; myopia.
Conflict of interest statement
All authors certify that they have no affiliations with or involvement in any organization or entity with any financial or non-financial interest in the subject matter or the materials discussed in this manuscript. The funders had no role in the design of the study; in the collection, analysis, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures






References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
