

N4-acetylcytidine of Nop2 mRNA is required for the transition of morula-to-blastocyst
- PMID: 37768430
- PMCID: PMC11071819
- DOI: 10.1007/s00018-023-04955-w
N4-acetylcytidine of Nop2 mRNA is required for the transition of morula-to-blastocyst
Abstract
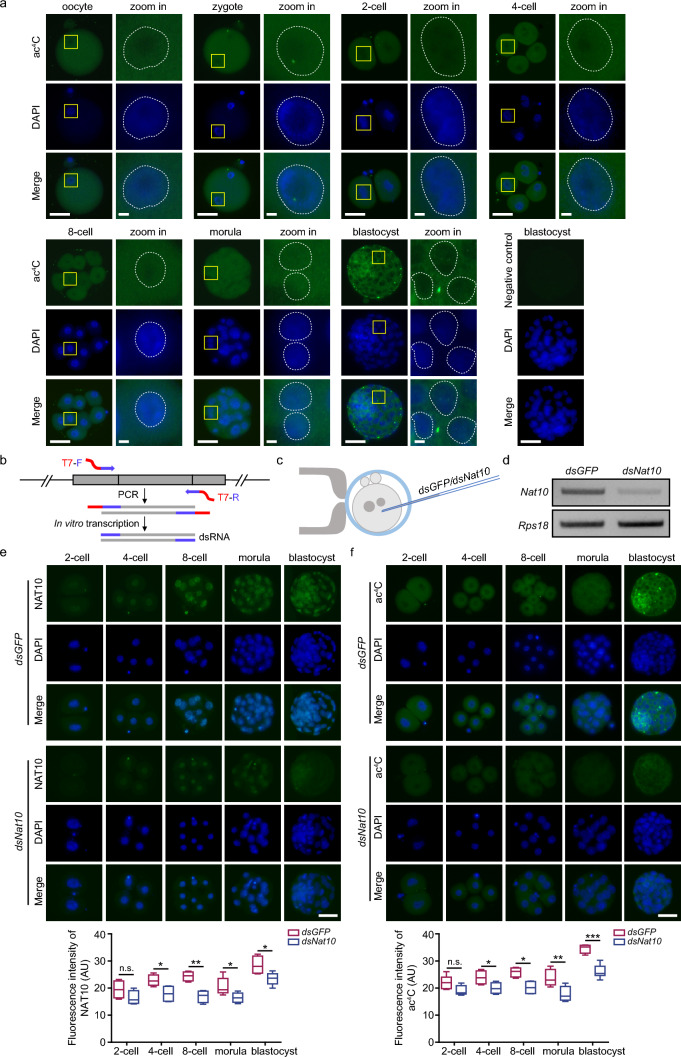
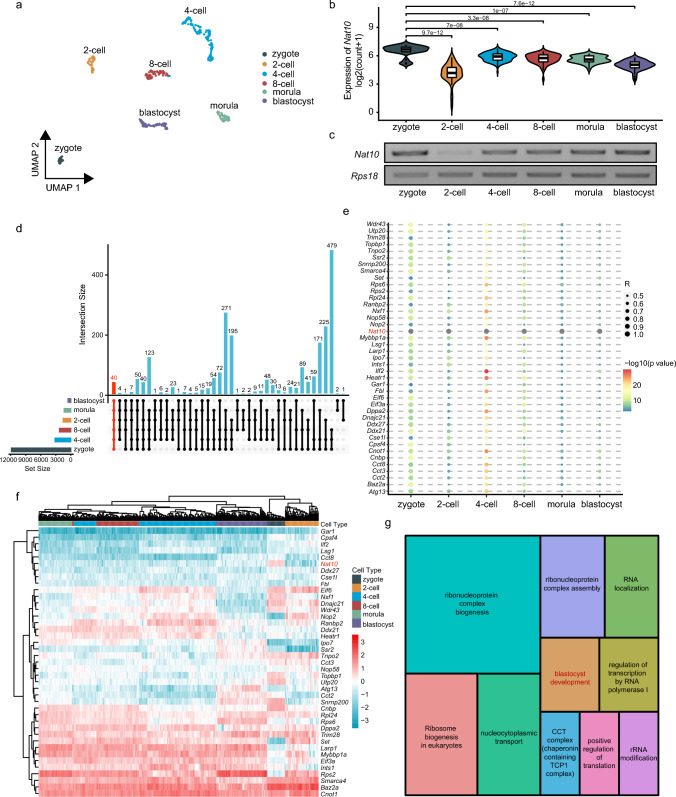
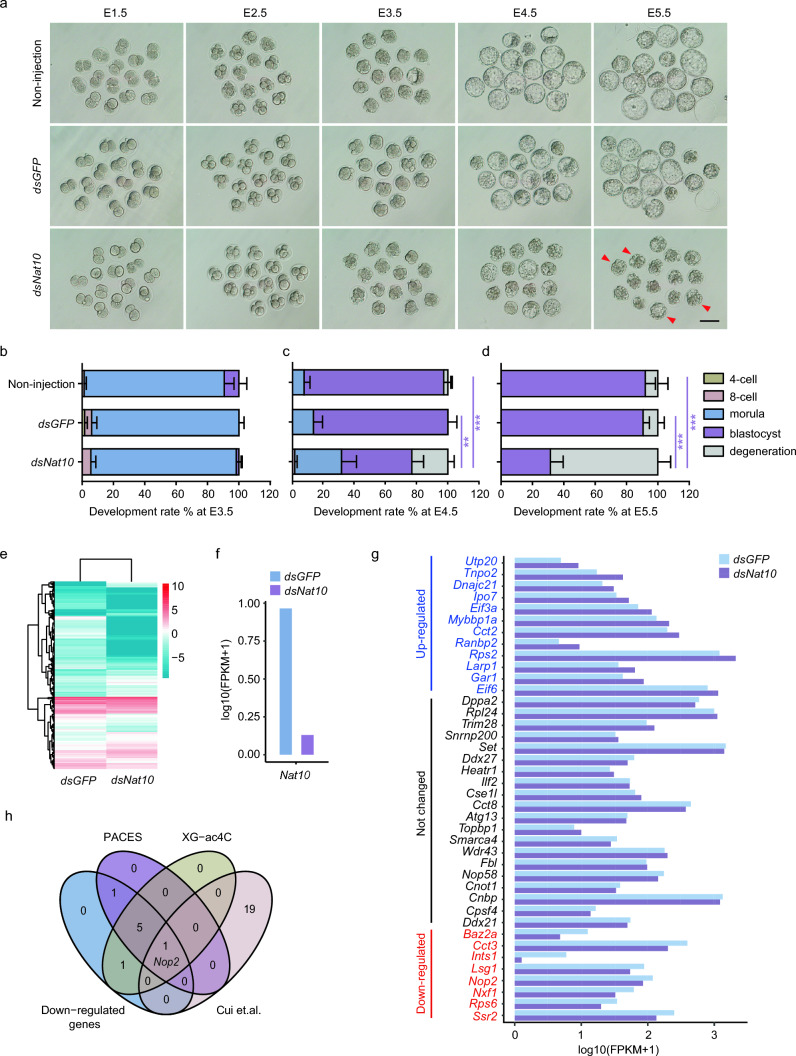
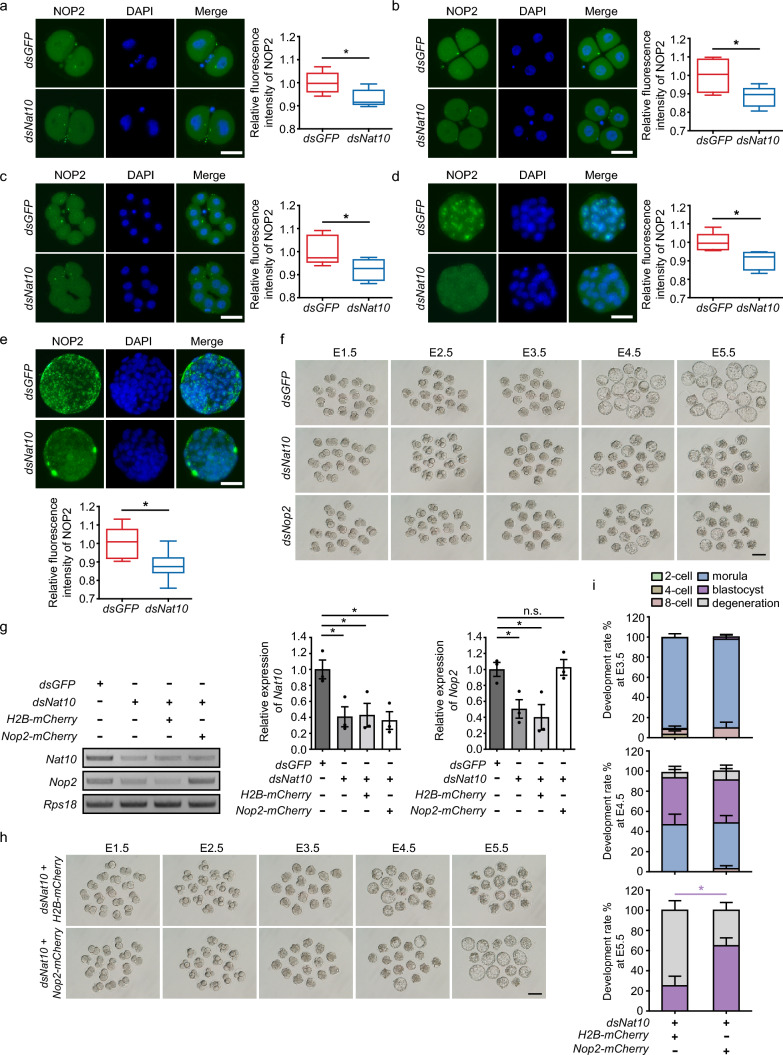
N-acetyltransferase 10 (NAT10)-mediated N4-acetylcytidine (ac4C) modification is crucial for mRNA stability and translation efficiency, yet the underlying function in mammalian preimplantation embryos remains unclear. Here, we characterized the ac4C modification landscape in mouse early embryos and found that the majority of embryos deficient in ac4C writer-NAT10 failed to develop into normal blastocysts. Through single-cell sequencing, RNA-seq, acetylated RNA immunoprecipitation combined with PCR (acRIP-PCR), and embryonic phenotype monitoring, Nop2 was screened as a target gene of Nat10. Mechanistically, Nat10 knockdown decreases the ac4C modification on Nop2 mRNA and reduces RNA and protein abundance by affecting the mRNA stability of Nop2. Then, depletion of NOP2 may inhibit the translation of transcription factor TEAD4, resulting in defective expression of the downstream lineage-specific gene Cdx2, and ultimately preventing blastomeres from undergoing the trophectoderm (TE) fate. However, exogenous Nop2 mRNA partially reverses this abnormal development. In conclusion, our findings demonstrate that defective ac4C modification of Nop2 mRNA hinders the morula-to-blastocyst transition by influencing the first cell fate decision in mice.
Keywords: Blastocyst formation; Development; Mouse; Preimplantation embryos; RNA modification.
© 2023. The Author(s), under exclusive licence to Springer Nature Switzerland AG.
Conflict of interest statement
The authors declare that they have no competing interests.
Figures



References
-
- Fleming TP, Pickering SJ. Maturation and polarization of the endocytotic system in outside blastomeres during mouse preimplantation development. J Embryol Exp Morphol. 1985;89:175–208. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
