

Structure and activation of the RING E3 ubiquitin ligase TRIM72 on the membrane
- PMID: 37770719
- PMCID: PMC10643145
- DOI: 10.1038/s41594-023-01111-7
Structure and activation of the RING E3 ubiquitin ligase TRIM72 on the membrane
Erratum in
-
Publisher Correction: Structure and activation of the RING E3 ubiquitin ligase TRIM72 on the membrane.Nat Struct Mol Biol. 2024 Nov;31(11):1810. doi: 10.1038/s41594-024-01429-w. Nat Struct Mol Biol. 2024. PMID: 39468339 Free PMC article. No abstract available.
Abstract
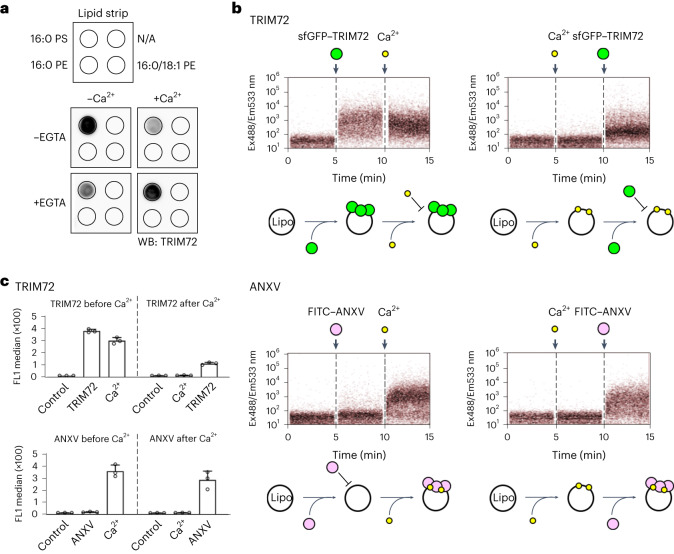
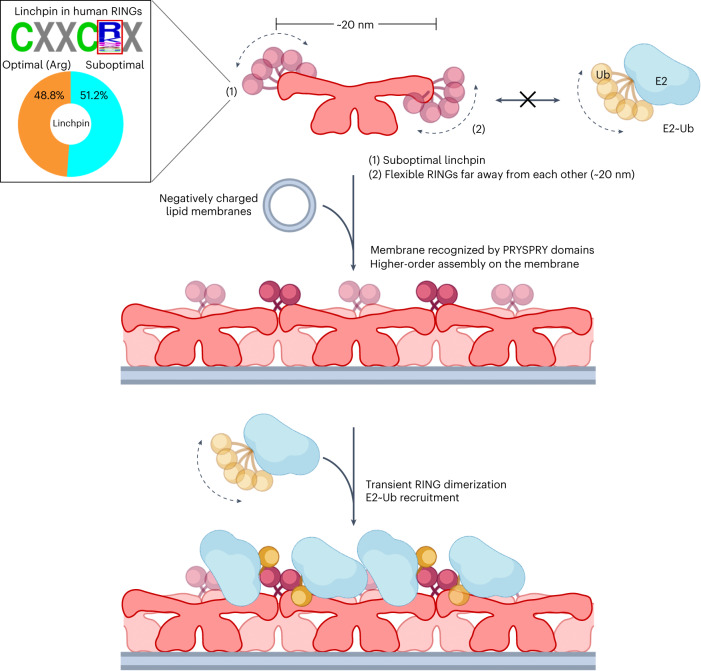
Defects in plasma membrane repair can lead to muscle and heart diseases in humans. Tripartite motif-containing protein (TRIM)72 (mitsugumin 53; MG53) has been determined to rapidly nucleate vesicles at the site of membrane damage, but the underlying molecular mechanisms remain poorly understood. Here we present the structure of Mus musculus TRIM72, a complete model of a TRIM E3 ubiquitin ligase. We demonstrated that the interaction between TRIM72 and phosphatidylserine-enriched membranes is necessary for its oligomeric assembly and ubiquitination activity. Using cryogenic electron tomography and subtomogram averaging, we elucidated a higher-order model of TRIM72 assembly on the phospholipid bilayer. Combining structural and biochemical techniques, we developed a working molecular model of TRIM72, providing insights into the regulation of RING-type E3 ligases through the cooperation of multiple domains in higher-order assemblies. Our findings establish a fundamental basis for the study of TRIM E3 ligases and have therapeutic implications for diseases associated with membrane repair.
© 2023. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures












References
-
- McNeil, P. L. & Kirchhausen, T. An emergency response team for membrane repair. Nat. Rev. Mol. Cell Biol.6, 499–505 (2005). - PubMed
-
- Liu, J. et al. Dysferlin, a novel skeletal muscle gene, is mutated in Miyoshi myopathy and limb girdle muscular dystrophy. Nat. Genet.20, 31–36 (1998). - PubMed
-
- Sun, M. et al. Mucolipidosis type IV is caused by mutations in a gene encoding a novel transient receptor potential channel. Hum. Mol. Genet.9, 2471–2478 (2000). - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
