

Oxamate enhances the efficacy of CAR-T therapy against glioblastoma via suppressing ectonucleotidases and CCR8 lactylation
- PMID: 37770937
- PMCID: PMC10540361
- DOI: 10.1186/s13046-023-02815-w
Oxamate enhances the efficacy of CAR-T therapy against glioblastoma via suppressing ectonucleotidases and CCR8 lactylation
Abstract
Background: Chimeric antigen receptor (CAR)-T immunotherapy fails to treat solid tumors due in part to immunosuppressive microenvironment. Excess lactate produced by tumor glycolysis increases CAR-T immunosuppression. The mechanism of lactate inducing the formation of immunosuppressive microenvironment remains to be further explored.
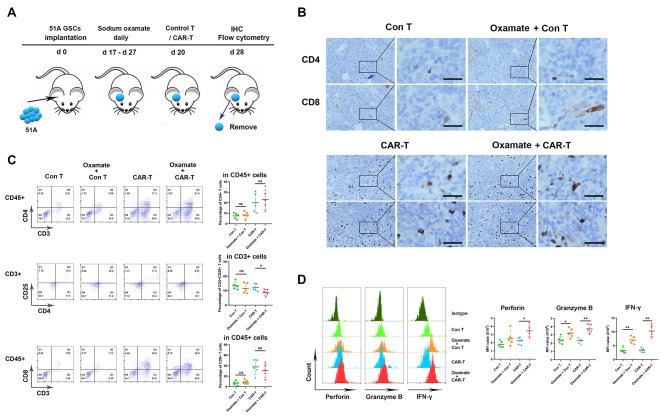
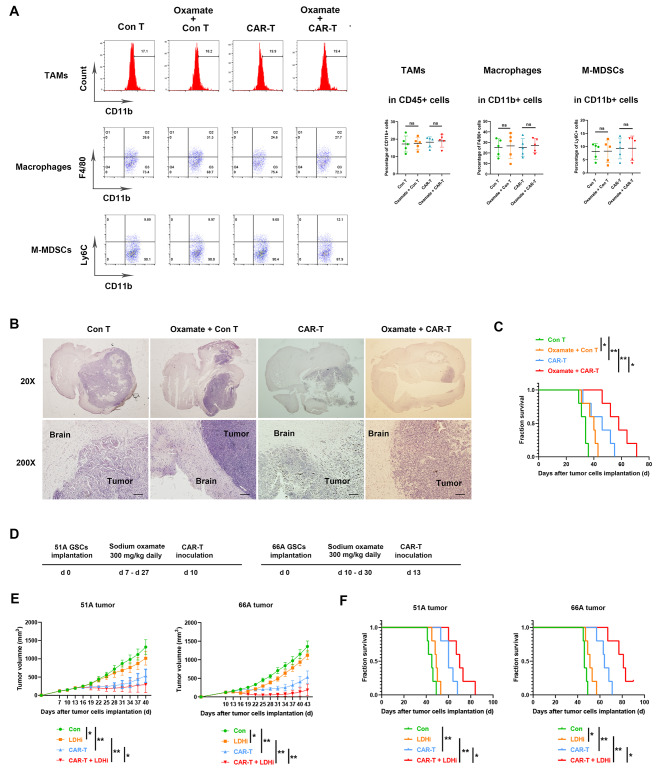
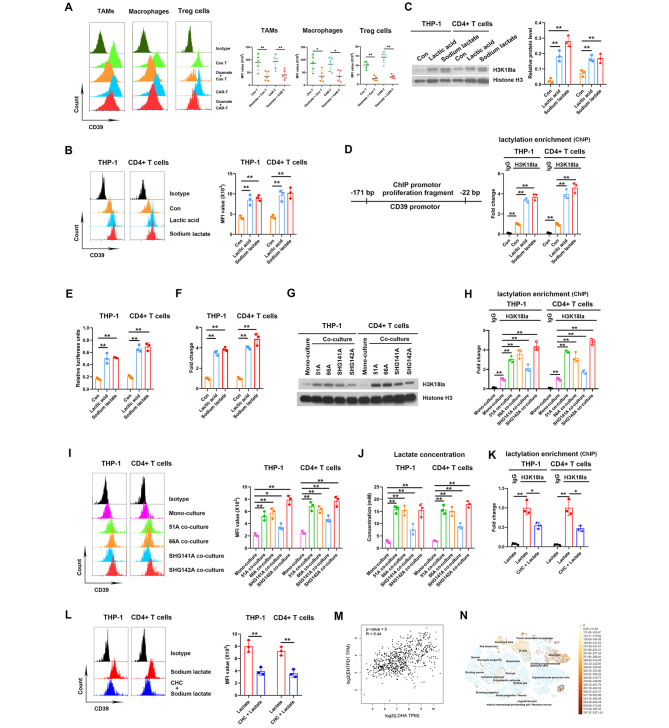
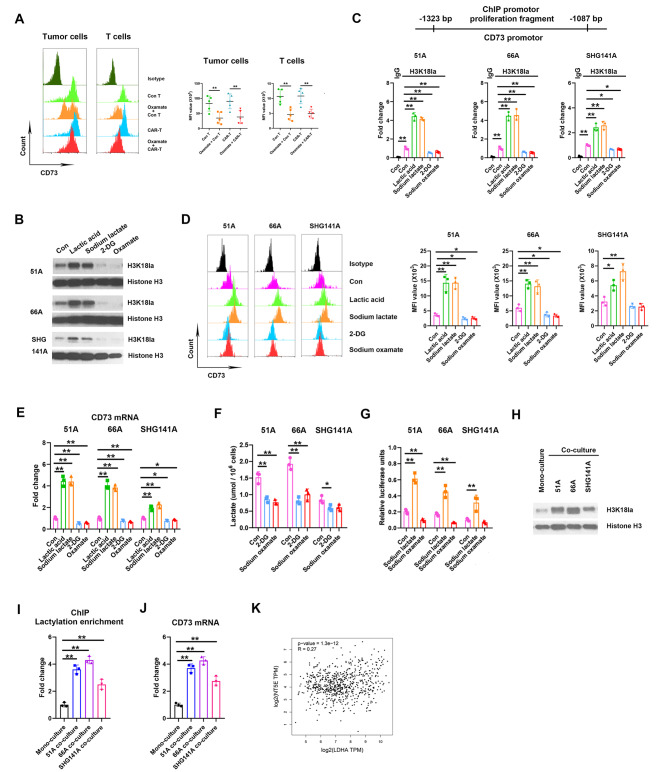
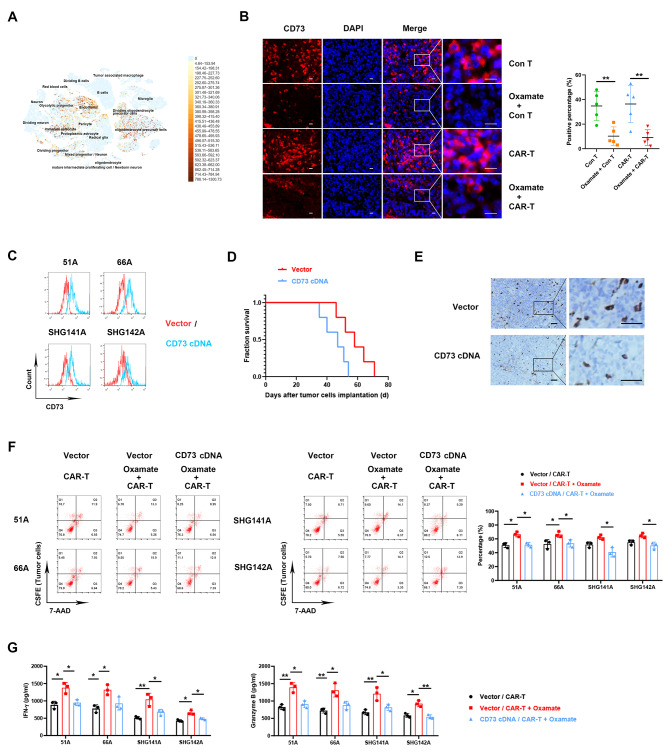
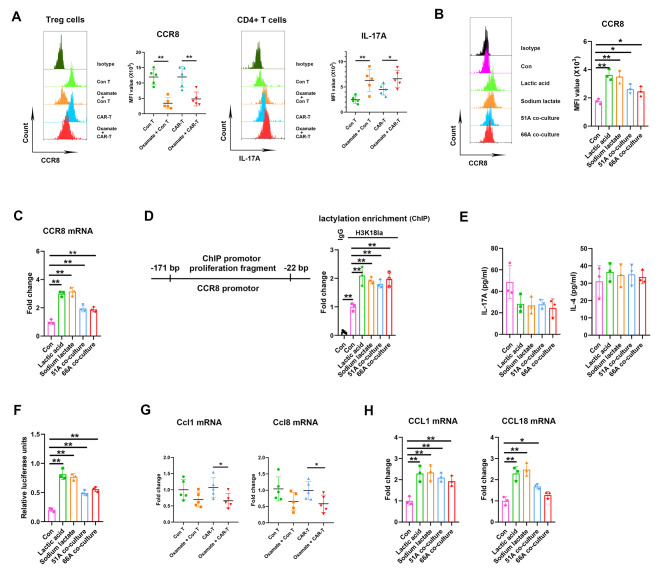
Methods: Immunocyte subpopulations and molecular characteristics were analyzed in the orthotopic xenografts of nude mice using flow cytometry assay and immunohistochemical staining after oxamate, a lactate dehydrogenase A (LDHA) inhibitor, and control T or CAR-T cells injection alone or in combination. RT-qPCR, western blot, flow cytometry, immunofluorescence, luciferase reporter assay, chromatin immunoprecipitation and ELISA were performed to measure the effect of lactate on the regulation of CD39, CD73 and CCR8 in cultured glioma stem cells, CD4 + T cells or macrophages.
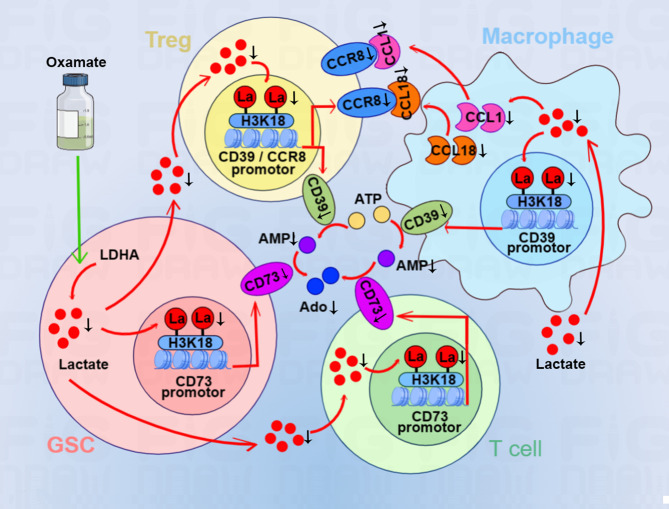
Results: Oxamate promoted immune activation of tumor-infiltrating CAR-T cells through altering the phenotypes of immune molecules and increasing regulatory T (Treg) cells infiltration in a glioblastoma mouse model. Lactate accumulation within cells upregulated CD39, CD73 and CCR8 expressions in both lactate-treated cells and glioma stem cells-co-cultured CD4 + T cells and macrophages, and intracellular lactate directly elevated the activities of these gene promotors through histone H3K18 lactylation.
Conclusions: Utilizing lactate generation inhibitor not only reprogramed glucose metabolism of cancer stem cells, but also alleviated immunosuppression of tumor microenvironment and reduced tumor-infiltrating CAR-Treg cells, which may be a potential strategy to enhance CAR-T function in glioblastoma therapy.
Keywords: CAR-T; CCR8; Ectonucleotidases; Glioblastoma; Histone lactylation.
© 2023. Italian National Cancer Institute ‘Regina Elena’.
Conflict of interest statement
No potential conflicts of interest were evident or relevant in the context of this article, and thus there was nothing to disclose.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
