

Markov chains as a proxy for the predictive memory representations underlying mismatch negativity
- PMID: 37771348
- PMCID: PMC10525344
- DOI: 10.3389/fnhum.2023.1249413
Markov chains as a proxy for the predictive memory representations underlying mismatch negativity
Abstract
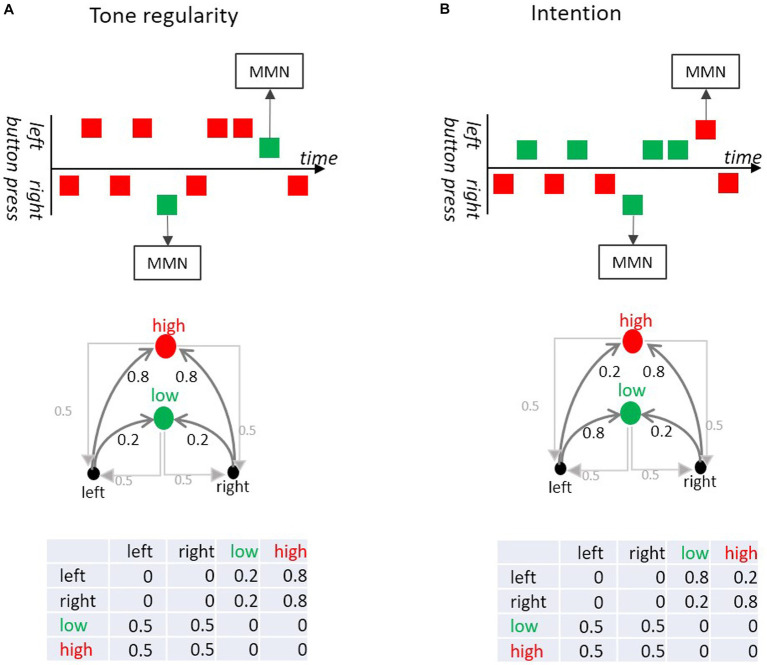
Events not conforming to a regularity inherent to a sequence of events elicit prediction error signals of the brain such as the Mismatch Negativity (MMN) and impair behavioral task performance. Events conforming to a regularity lead to attenuation of brain activity such as stimulus-specific adaptation (SSA) and behavioral benefits. Such findings are usually explained by theories stating that the information processing system predicts the forthcoming event of the sequence via detected sequential regularities. A mathematical model that is widely used to describe, to analyze and to generate event sequences are Markov chains: They contain a set of possible events and a set of probabilities for transitions between these events (transition matrix) that allow to predict the next event on the basis of the current event and the transition probabilities. The accuracy of such a prediction depends on the distribution of the transition probabilities. We argue that Markov chains also have useful applications when studying cognitive brain functions. The transition matrix can be regarded as a proxy for generative memory representations that the brain uses to predict the next event. We assume that detected regularities in a sequence of events correspond to (a subset of) the entries in the transition matrix. We apply this idea to the Mismatch Negativity (MMN) research and examine three types of MMN paradigms: classical oddball paradigms emphasizing sound probabilities, between-sound regularity paradigms manipulating transition probabilities between adjacent sounds, and action-sound coupling paradigms in which sounds are associated with actions and their intended effects. We show that the Markovian view on MMN yields theoretically relevant insights into the brain processes underlying MMN and stimulates experimental designs to study the brain's processing of event sequences.
Keywords: Markov model; adaptation; memory; mismatch negativity; perception; predictive processing; regularity representation.
Copyright © 2023 Schröger, Roeber and Coy.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






References
LinkOut - more resources
Full Text Sources
Research Materials