

An autoinhibited state of 53BP1 revealed by small molecule antagonists and protein engineering
- PMID: 37773238
- PMCID: PMC10541411
- DOI: 10.1038/s41467-023-41821-6
An autoinhibited state of 53BP1 revealed by small molecule antagonists and protein engineering
Abstract
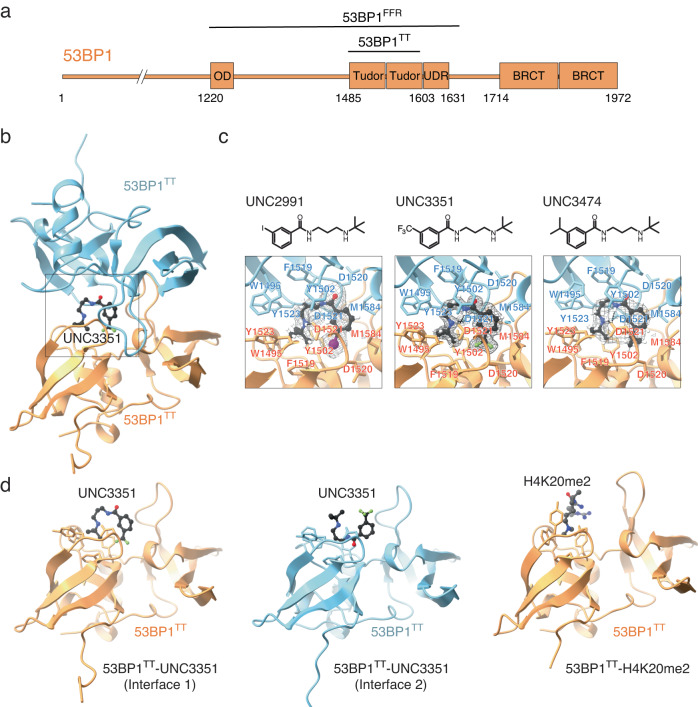
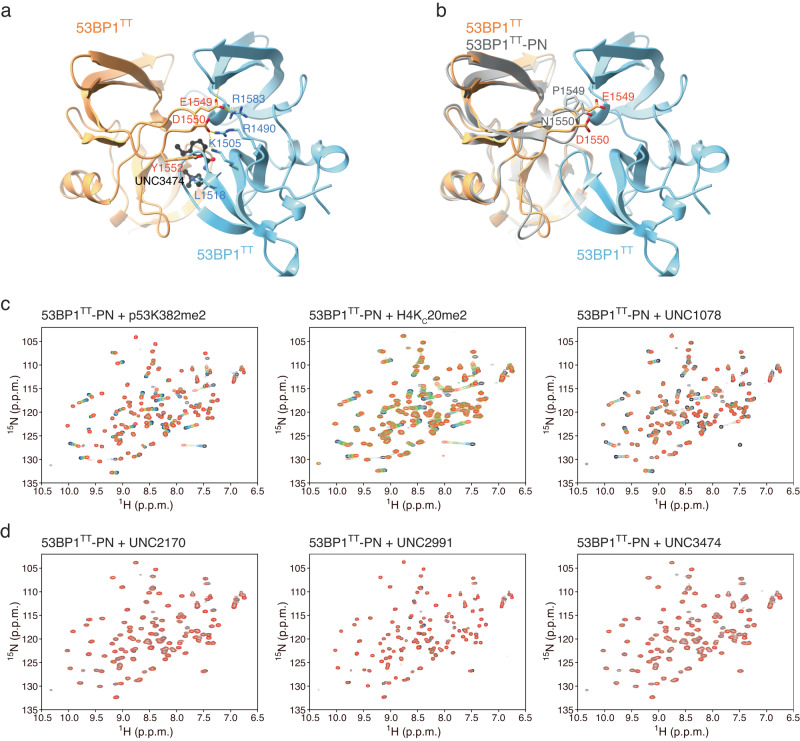
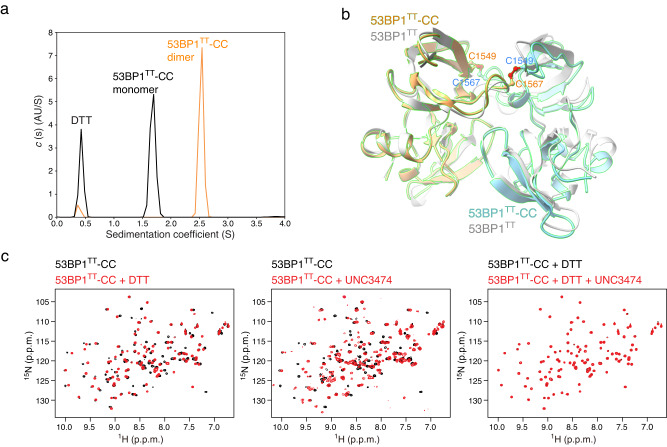
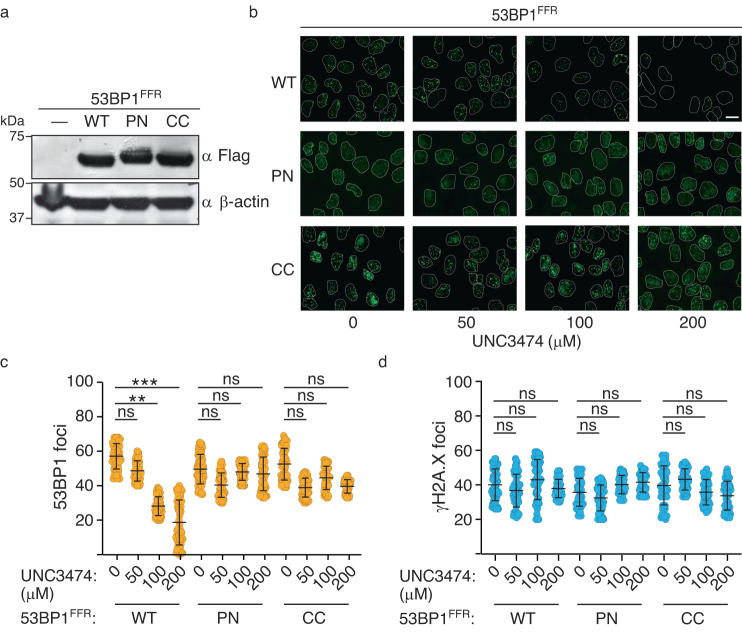
The recruitment of 53BP1 to chromatin, mediated by its recognition of histone H4 dimethylated at lysine 20 (H4K20me2), is important for DNA double-strand break repair. Using a series of small molecule antagonists, we demonstrate a conformational equilibrium between an open and a pre-existing lowly populated closed state of 53BP1 in which the H4K20me2 binding surface is buried at the interface between two interacting 53BP1 molecules. In cells, these antagonists inhibit the chromatin recruitment of wild type 53BP1, but do not affect 53BP1 variants unable to access the closed conformation despite preservation of the H4K20me2 binding site. Thus, this inhibition operates by shifting the conformational equilibrium toward the closed state. Our work therefore identifies an auto-associated form of 53BP1-autoinhibited for chromatin binding-that can be stabilized by small molecule ligands encapsulated between two 53BP1 protomers. Such ligands are valuable research tools to study the function of 53BP1 and have the potential to facilitate the development of new drugs for cancer therapy.
© 2023. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures

Update of
-
An autoinhibited state of 53BP1 revealed by small molecule antagonists and protein engineering.bioRxiv [Preprint]. 2023 Jul 18:2023.04.20.534960. doi: 10.1101/2023.04.20.534960. bioRxiv. 2023. Update in: Nat Commun. 2023 Sep 29;14(1):6091. doi: 10.1038/s41467-023-41821-6. PMID: 37131705 Free PMC article. Updated. Preprint.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
